XiaoMi-AI文件搜索系统
World File Search System类癌

肿瘤微环境相互作用引导图形学习,从全切片病理图像进行人类癌症生存分析
深度学习技术的最新进展为协助病理学家从全切片病理图像(WSI)中预测患者的生存期带来了可能性。然而,大多数流行的方法仅适用于WSI中特定或随机选择的肿瘤区域中的采样斑块,这对于捕捉肿瘤与其周围微环境成分之间复杂相互作用的能力非常有限。事实上,肿瘤在异质性肿瘤微环境(TME)中得到支持和培育,详细分析TME及其与肿瘤的相关性对于深入分析癌症发展的机制具有重要意义。在本文中,我们考虑了肿瘤与其两个主要TME成分(即淋巴细胞和基质纤维化)之间的空间相互作用,并提出了一种用于人类癌症预后预测的肿瘤微环境相互作用引导图学习(TMEGL)算法。具体来说,我们首先选择不同类型的块作为节点来为每个 WSI 构建图。然后,提出了一种新颖的 TME 邻域组织引导图嵌入算法来学习可以保留其拓扑结构信息的节点表示。最后,应用门控图注意网络来捕获肿瘤与不同 TME 组件之间与生存相关的交集以进行临床结果预测。我们在来自癌症基因组图谱 (TCGA) 的三个癌症队列上测试了 TMEGL,实验结果表明 TMEGL 不仅优于现有的基于 WSI 的生存分析模型,而且对生存预测具有良好的可解释能力。

利用功能基因组学和人工智能对人类癌细胞进行逆向工程
我写这本书的首要动机是一句您将在接下来的内容中多次看到的短语。这句话是:“转移性疾病无法治愈”。这句话之所以如此重要,是因为尽管我们在癌症研究方面取得了数十年的巨大进步,但一旦疾病扩散到远处器官,患者的治疗进展就非常有限。正因为如此,我们作为一个社区显然是时候尝试一些新方法了,因为标准化疗虽然在疾病的其他阶段有用,但无法让我们到达最后的顶峰,即转移性癌症的治愈。在我看来,其中一种这样的策略涉及将现代人工智能 (AI) 和机器学习 (ML) 方法应用于从癌症患者和癌症衍生细胞系中积累的大量基因组数据,以制定真正个性化的策略,以对个体患者进行癌症逆向工程。因此,本书的目标是让读者相信这是可能的,至少是一条值得追求的途径。首先我要说的是,我将在本书中强调人工智能对基因组数据的分析如何帮助我们更好地利用癌症靶向疗法。与此同时,其他人也在努力开发类似的方法,利用计算和人工智能方法来改善癌症免疫疗法的使用,因为免疫疗法提供了另一套可用于转移性癌症患者的工具。由于我不是免疫学家,我不会在这里讨论这些方法,因为它们可以在其他出版物中找到。

生发中心依赖性和 - 非依赖性的免疫反应在人类癌症中肿瘤的B细胞
b细胞在免疫中起着重要作用,主要是通过产生高质量浆细胞(PC)和记忆B(BMEM)细胞。分别依赖于抗原结合和微环境提供的B细胞受体(BCR)固有和外在信号的B细胞(BCR)固有和外在信号的整合。近年来,滤觉B(TIL-B)细胞(TIL-B)和PC(TIL-PC)中的肿瘤已被揭示为人类癌症中抗肿瘤反应的重要参与者,但是它们的相互作用和动态仍然很少知道。在淋巴机构中,B细胞反应涉及BMEM细胞和PC产生的生发中心(GC)依赖性和与GC独立的途径。affiential bcr库的成熟发生在GC反应中,具有B细胞信号积分的特定时空动力学。通常,抗原通过抗原触发GC独立于产生大量PC而无需BCR重生的抗原的生产。了解免疫反应中的B细胞动力学需要多种工具和读数(例如单细胞表型和RNA-SEQ),原位分析,BCR曲目分析,BCR特异性和依次范围的fifirity分析和功能测试和功能测试。在这里,我们回顾了如何将这些工具应用于不同类型的实体瘤中的TIL-B细胞和TIL-PC。我们评估了涉及涉及GC依赖性或独立于GC的局部响应的TIL-B细胞动力学不同模型的已公开证据,以及由抗原特异性PC的产生。总的来说,我们强调了需要进行更整合的B细胞免疫学研究,以合理研究TIL-B细胞作为抗肿瘤疗法的杠杆作用。

作者校正:一个多线屏幕确定人类癌症中可起作的合成致命相互作用
Open Access本文在创意共享属性 - 非商业性 - 发病率4.0国际许可下获得许可,该许可允许以任何中等或格式的任何非商业用途,共享,分发和复制,只要您与原始作者提供适当的信誉,并提供了与创意共享许可证的链接,如果您提供了持牌材料,并提供了启动的链接。您没有根据本许可证的许可来共享本文或部分内容的适用材料。本文中的图像或其他第三方材料包含在文章的创意共享许可中,除非在信用额度中另有说明。如果本文的创意共享许可中未包含材料,并且您的预期用途不受法定法规的允许或超过允许的用途,则您需要直接从版权所有者那里获得许可。要查看此许可证的副本,请访问http://creativecommons.org/licenses/by-nc-nd/4.0/。
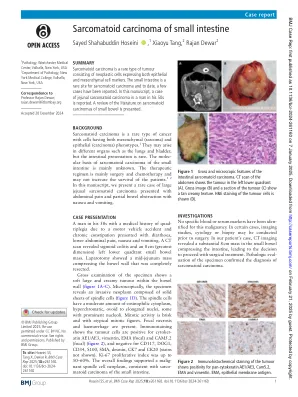
小肠的肉瘤类癌
案例表现,一名50多岁的男人患有四肢际病史,这是由于汽车事故和慢性便秘,腹泻,下腹痛,恶心和呕吐。CT扫描显示乙状结肠炎和8厘米(最大维度)左下象限小肠质量。剖腹手术显示出完全切除的肠壁中的jejunum质量。对试样的总检查显示了肠壁内柔软的大型乳脂状肿瘤(图1A – C)。显微镜下,样品揭示了由纺锤体细胞实心板组成的侵入性肿瘤(图1D)。纺锤体细胞具有适量的嗜酸性细胞质,过度骨质,卵形对细长核,有些具有突出的核仁。有丝分裂活性是轻快的,具有非典型有丝分裂数字。存在局灶性坏死和出血。免疫染色表明肿瘤细胞的阳性是阳性的AE1/AE3,Vimentin,Ema(焦点)和CAM5.2(焦点)(图2),而CD117,DOG1,CD34,S100,S100,SMA,Desmin,desmin,ck7和ck20(未显示)(未显示)。KI-67增殖指数高达50–60%。 总体发现支持了与小肠的肉眼癌癌相一致的杂质纺锤体肿瘤。KI-67增殖指数高达50–60%。总体发现支持了与小肠的肉眼癌癌相一致的杂质纺锤体肿瘤。
![b'[2] C. Yan,X。Duanmu,L。Zeng,B。Liu,Z。Song,线粒体DNA:分布,突变和消除,细胞,8(2019)。 [3] F. Liu,D.E。 Sanin,X。Wang,肺癌中的线粒体DNA,实验医学和生物学进展,1038(2017)9-22。 [4] J. Zhang,J。 Jia,M。Junaid,Y.B。 MA,F。Ahmad,Y.F。 Jia,W.G。Li,D.S。Pei,人类DNA2(HDNA2)作为癌症和其他疾病的潜在靶标的作用:系统评价,DNA修复(AMST),59(2017)9-19。 [6] A. D \ XC3 \ Xadaz-Talavera,C。Montero-Conde,L.J。Leandro-Garc \ XC3 \ XCAD \ Xada,M。Robledo,Primpol:DNA复制酶之间的突破性和潜在的癌症治疗靶标的癌症治疗,生物分子,12(20222)。 [7] A.O. Giacomelli,X。Yang,R.E。 Lintner,J.M。McFarland,M。Duby,J.Kim,T.P。 D.Y. Howard武田Ly,E。Kim,H.S。甘农(B. J.G. Aguirre Doench,F。Piccioni,C.W.M。 Roberts,M。Meyerson,G。Getz,C.M。约翰内森,D.E。根,W.C。 Hahn,突变过程塑造了人类癌症中TP53突变的景观,NAT Genet,50(2018)1381-1387。 [8] G.A. Fontana,H.L。Gahlon,线粒体DNA缺失形成的复制和修复机制,核酸res,48(2020)11244-11258。 [9] C.Y. Dai,C.C。 Ng,G.C.C。 Hung,I。Kirmes,L.A](/simg/f/f6a2bb5f504472f4ca10c1ecebfe7702878cdb55.webp)
b'[2] C. Yan,X。Duanmu,L。Zeng,B。Liu,Z。Song,线粒体DNA:分布,突变和消除,细胞,8(2019)。 [3] F. Liu,D.E。 Sanin,X。Wang,肺癌中的线粒体DNA,实验医学和生物学进展,1038(2017)9-22。 [4] J. Zhang,J。 Jia,M。Junaid,Y.B。 MA,F。Ahmad,Y.F。 Jia,W.G。Li,D.S。Pei,人类DNA2(HDNA2)作为癌症和其他疾病的潜在靶标的作用:系统评价,DNA修复(AMST),59(2017)9-19。 [6] A. D \ XC3 \ Xadaz-Talavera,C。Montero-Conde,L.J。Leandro-Garc \ XC3 \ XCAD \ Xada,M。Robledo,Primpol:DNA复制酶之间的突破性和潜在的癌症治疗靶标的癌症治疗,生物分子,12(20222)。 [7] A.O. Giacomelli,X。Yang,R.E。 Lintner,J.M。McFarland,M。Duby,J.Kim,T.P。 D.Y. Howard武田Ly,E。Kim,H.S。甘农(B. J.G. Aguirre Doench,F。Piccioni,C.W.M。 Roberts,M。Meyerson,G。Getz,C.M。约翰内森,D.E。根,W.C。 Hahn,突变过程塑造了人类癌症中TP53突变的景观,NAT Genet,50(2018)1381-1387。 [8] G.A. Fontana,H.L。Gahlon,线粒体DNA缺失形成的复制和修复机制,核酸res,48(2020)11244-11258。 [9] C.Y. Dai,C.C。 Ng,G.C.C。 Hung,I。Kirmes,L.A
b'[2] C. Yan,X。Duanmu,L。Zeng,B。Liu,Z。歌曲,线粒体DNA:分布,突变和消除,细胞,8(2019)。[3] F. Liu,D.E。Sanin,X。Wang,肺癌中的线粒体DNA,实验医学与生物学进展,1038(2017)9-22。[4] J. Zhang,J。[5] P.P.Jia,M。Junaid,Y.B。 MA,F。Ahmad,Y.F。 jia,W.G。 li,D.S。 pei,人类DNA2(HDNA2)作为癌症和其他疾病的潜在靶点的作用:系统评价,DNA修复(AMST),59(2017)9-19。 [6] A. D \ XC3 \ Xadaz-Talavera,C。Montero-Conde,L.J。 Leandro-Garc \ XC3 \ Xada,M。Robledo,Primpol:DNA复制酶的突破和潜在的癌症治疗新靶标,生物分子,12(2022)。 [7] A.O. Giacomelli,X。Yang,R.E。 lintner,J.M. McFarland,M。Duby,J。Kim,T.P。 D.Y. Howard Takeda,S.H。 ly,E。Kim,H.S。 Gannon,B。Hurhula,T。Sharpe,A。Goodale,B。Fritchman,S。Seelman,F。Vazquez,A。Tsherniak,A.J。 Aguirre,J.G。 Doench,F。Piccioni,C.W.M。 Roberts,M。Meyerson,G。Getz,C.M。 Johannessen,D.E。 根,W.C。 Hahn,突变过程塑造了人类癌症中TP53突变的景观,NAT Genet,50(2018)1381-1387。 [8] G.A. Fontana,H.L。 [9] C.Y. dai,C.C。 ng,G.C.C。 Hung,I。Kirmes,L.A。Hughes,Y。Jia,M。Junaid,Y.B。MA,F。Ahmad,Y.F。 jia,W.G。 li,D.S。 pei,人类DNA2(HDNA2)作为癌症和其他疾病的潜在靶点的作用:系统评价,DNA修复(AMST),59(2017)9-19。 [6] A. D \ XC3 \ Xadaz-Talavera,C。Montero-Conde,L.J。 Leandro-Garc \ XC3 \ Xada,M。Robledo,Primpol:DNA复制酶的突破和潜在的癌症治疗新靶标,生物分子,12(2022)。 [7] A.O. Giacomelli,X。Yang,R.E。 lintner,J.M. McFarland,M。Duby,J。Kim,T.P。 D.Y. Howard Takeda,S.H。 ly,E。Kim,H.S。 Gannon,B。Hurhula,T。Sharpe,A。Goodale,B。Fritchman,S。Seelman,F。Vazquez,A。Tsherniak,A.J。 Aguirre,J.G。 Doench,F。Piccioni,C.W.M。 Roberts,M。Meyerson,G。Getz,C.M。 Johannessen,D.E。 根,W.C。 Hahn,突变过程塑造了人类癌症中TP53突变的景观,NAT Genet,50(2018)1381-1387。 [8] G.A. Fontana,H.L。 [9] C.Y. dai,C.C。 ng,G.C.C。 Hung,I。Kirmes,L.A。Hughes,Y。MA,F。Ahmad,Y.F。jia,W.G。li,D.S。pei,人类DNA2(HDNA2)作为癌症和其他疾病的潜在靶点的作用:系统评价,DNA修复(AMST),59(2017)9-19。[6] A. D \ XC3 \ Xadaz-Talavera,C。Montero-Conde,L.J。Leandro-Garc \ XC3 \ Xada,M。Robledo,Primpol:DNA复制酶的突破和潜在的癌症治疗新靶标,生物分子,12(2022)。[7] A.O.Giacomelli,X。Yang,R.E。 lintner,J.M. McFarland,M。Duby,J。Kim,T.P。 D.Y. Howard Takeda,S.H。 ly,E。Kim,H.S。 Gannon,B。Hurhula,T。Sharpe,A。Goodale,B。Fritchman,S。Seelman,F。Vazquez,A。Tsherniak,A.J。 Aguirre,J.G。 Doench,F。Piccioni,C.W.M。 Roberts,M。Meyerson,G。Getz,C.M。 Johannessen,D.E。 根,W.C。 Hahn,突变过程塑造了人类癌症中TP53突变的景观,NAT Genet,50(2018)1381-1387。 [8] G.A. Fontana,H.L。 [9] C.Y. dai,C.C。 ng,G.C.C。 Hung,I。Kirmes,L.A。Hughes,Y。Giacomelli,X。Yang,R.E。lintner,J.M.McFarland,M。Duby,J。Kim,T.P。 D.Y. Howard Takeda,S.H。 ly,E。Kim,H.S。 Gannon,B。Hurhula,T。Sharpe,A。Goodale,B。Fritchman,S。Seelman,F。Vazquez,A。Tsherniak,A.J。 Aguirre,J.G。 Doench,F。Piccioni,C.W.M。 Roberts,M。Meyerson,G。Getz,C.M。 Johannessen,D.E。 根,W.C。 Hahn,突变过程塑造了人类癌症中TP53突变的景观,NAT Genet,50(2018)1381-1387。 [8] G.A. Fontana,H.L。 [9] C.Y. dai,C.C。 ng,G.C.C。 Hung,I。Kirmes,L.A。Hughes,Y。McFarland,M。Duby,J。Kim,T.P。D.Y. Howard Takeda,S.H。 ly,E。Kim,H.S。 Gannon,B。Hurhula,T。Sharpe,A。Goodale,B。Fritchman,S。Seelman,F。Vazquez,A。Tsherniak,A.J。 Aguirre,J.G。 Doench,F。Piccioni,C.W.M。 Roberts,M。Meyerson,G。Getz,C.M。 Johannessen,D.E。 根,W.C。 Hahn,突变过程塑造了人类癌症中TP53突变的景观,NAT Genet,50(2018)1381-1387。 [8] G.A. Fontana,H.L。 [9] C.Y. dai,C.C。 ng,G.C.C。 Hung,I。Kirmes,L.A。Hughes,Y。D.Y. HowardTakeda,S.H。 ly,E。Kim,H.S。 Gannon,B。Hurhula,T。Sharpe,A。Goodale,B。Fritchman,S。Seelman,F。Vazquez,A。Tsherniak,A.J。 Aguirre,J.G。 Doench,F。Piccioni,C.W.M。 Roberts,M。Meyerson,G。Getz,C.M。 Johannessen,D.E。 根,W.C。 Hahn,突变过程塑造了人类癌症中TP53突变的景观,NAT Genet,50(2018)1381-1387。 [8] G.A. Fontana,H.L。 [9] C.Y. dai,C.C。 ng,G.C.C。 Hung,I。Kirmes,L.A。Hughes,Y。Takeda,S.H。ly,E。Kim,H.S。Gannon,B。Hurhula,T。Sharpe,A。Goodale,B。Fritchman,S。Seelman,F。Vazquez,A。Tsherniak,A.J。 Aguirre,J.G。 Doench,F。Piccioni,C.W.M。 Roberts,M。Meyerson,G。Getz,C.M。 Johannessen,D.E。 根,W.C。 Hahn,突变过程塑造了人类癌症中TP53突变的景观,NAT Genet,50(2018)1381-1387。 [8] G.A. Fontana,H.L。 [9] C.Y. dai,C.C。 ng,G.C.C。 Hung,I。Kirmes,L.A。Hughes,Y。Gannon,B。Hurhula,T。Sharpe,A。Goodale,B。Fritchman,S。Seelman,F。Vazquez,A。Tsherniak,A.J。Aguirre,J.G。 Doench,F。Piccioni,C.W.M。 Roberts,M。Meyerson,G。Getz,C.M。 Johannessen,D.E。 根,W.C。 Hahn,突变过程塑造了人类癌症中TP53突变的景观,NAT Genet,50(2018)1381-1387。 [8] G.A. Fontana,H.L。 [9] C.Y. dai,C.C。 ng,G.C.C。 Hung,I。Kirmes,L.A。Hughes,Y。Aguirre,J.G。Doench,F。Piccioni,C.W.M。Roberts,M。Meyerson,G。Getz,C.M。 Johannessen,D.E。 根,W.C。 Hahn,突变过程塑造了人类癌症中TP53突变的景观,NAT Genet,50(2018)1381-1387。 [8] G.A. Fontana,H.L。 [9] C.Y. dai,C.C。 ng,G.C.C。 Hung,I。Kirmes,L.A。Hughes,Y。Roberts,M。Meyerson,G。Getz,C.M。Johannessen,D.E。 根,W.C。 Hahn,突变过程塑造了人类癌症中TP53突变的景观,NAT Genet,50(2018)1381-1387。 [8] G.A. Fontana,H.L。 [9] C.Y. dai,C.C。 ng,G.C.C。 Hung,I。Kirmes,L.A。Hughes,Y。Johannessen,D.E。根,W.C。 Hahn,突变过程塑造了人类癌症中TP53突变的景观,NAT Genet,50(2018)1381-1387。[8] G.A.Fontana,H.L。 [9] C.Y. dai,C.C。 ng,G.C.C。 Hung,I。Kirmes,L.A。Hughes,Y。Fontana,H.L。[9] C.Y.dai,C.C。ng,G.C.C。Hung,I。Kirmes,L.A。Hughes,Y。gahlon,线粒体DNA缺失形成的复制和修复机制,核酸res,48(2020)11244-11258。du,C.A。Brosnan,A。Ahier,A。Hahn,C.M。 Haynes,O。Rackham,A。Filipovska,S。Zuryn,ATFS-1,通过促进转录修复,自然细胞生物学,25(2023)1111-1120来抵消线粒体DNA损伤。 [10] L. Ou,H。Liu,C。Peng,Y. [11] H. Liu,J。Weng,C.L.H。 Huang,A.P。 杰克逊,癌症的电压门控钠通道,生物标志物研究,12(2024)70。 [12] H. Liu,A。Dong,A.M。 Rasteh,P。Wang,J。Weng,乳腺癌中新型T细胞CD8 +标记的鉴定,Scientific Reports,14(2024)19142。 [13] H. Liu,T。Tang,基于MAPK信号途径的胶质瘤亚型,机器学习风险模型和关键集线器蛋白识别,科学报告,13(2023)19055。。 [14] H. Liu,T。Tang,《泛滥成灾与基因集的泛癌遗传分析》,癌症遗传学,278-279(2023)91-103。 [15] H. Liu,T。Tang,《胶质瘤IGFBP的生物信息学研究》,涉及其诊断,预后和治疗预测值,AM J Transl Res,15(2023)2140-2155。 [16] H. Liu,T。Tang,《泛滥成灾基因套件的泛 - 癌遗传分析》,Biorxiv,(2023),2023.2002。 2025.529997。 [17] H. Liu,库糖凋亡在肾脏肾透明细胞癌中的表达和潜在免疫受累,癌症遗传学,274-275(2023)21-25。Brosnan,A。Ahier,A。Hahn,C.M。Haynes,O。Rackham,A。Filipovska,S。Zuryn,ATFS-1,通过促进转录修复,自然细胞生物学,25(2023)1111-1120来抵消线粒体DNA损伤。[10] L. Ou,H。Liu,C。Peng,Y.[11] H. Liu,J。Weng,C.L.H。Huang,A.P。 杰克逊,癌症的电压门控钠通道,生物标志物研究,12(2024)70。 [12] H. Liu,A。Dong,A.M。 Rasteh,P。Wang,J。Weng,乳腺癌中新型T细胞CD8 +标记的鉴定,Scientific Reports,14(2024)19142。 [13] H. Liu,T。Tang,基于MAPK信号途径的胶质瘤亚型,机器学习风险模型和关键集线器蛋白识别,科学报告,13(2023)19055。。 [14] H. Liu,T。Tang,《泛滥成灾与基因集的泛癌遗传分析》,癌症遗传学,278-279(2023)91-103。 [15] H. Liu,T。Tang,《胶质瘤IGFBP的生物信息学研究》,涉及其诊断,预后和治疗预测值,AM J Transl Res,15(2023)2140-2155。 [16] H. Liu,T。Tang,《泛滥成灾基因套件的泛 - 癌遗传分析》,Biorxiv,(2023),2023.2002。 2025.529997。 [17] H. Liu,库糖凋亡在肾脏肾透明细胞癌中的表达和潜在免疫受累,癌症遗传学,274-275(2023)21-25。Huang,A.P。杰克逊,癌症的电压门控钠通道,生物标志物研究,12(2024)70。[12] H. Liu,A。Dong,A.M。 Rasteh,P。Wang,J。Weng,乳腺癌中新型T细胞CD8 +标记的鉴定,Scientific Reports,14(2024)19142。[13] H. Liu,T。Tang,基于MAPK信号途径的胶质瘤亚型,机器学习风险模型和关键集线器蛋白识别,科学报告,13(2023)19055。[14] H. Liu,T。Tang,《泛滥成灾与基因集的泛癌遗传分析》,癌症遗传学,278-279(2023)91-103。[15] H. Liu,T。Tang,《胶质瘤IGFBP的生物信息学研究》,涉及其诊断,预后和治疗预测值,AM J Transl Res,15(2023)2140-2155。[16] H. Liu,T。Tang,《泛滥成灾基因套件的泛 - 癌遗传分析》,Biorxiv,(2023),2023.2002。2025.529997。[17] H. Liu,库糖凋亡在肾脏肾透明细胞癌中的表达和潜在免疫受累,癌症遗传学,274-275(2023)21-25。[18] L. Hengrui,《中药用于癌症治疗中使用的有毒药物的例子》,J Tradit Chin Med,43(2023)209-210。[19] H. Liu,J。Weng,《 Rad51的Pan-Cancer生物信息学分析》,涉及诊断,预后和治疗预测的值,肿瘤学的前沿,12(2022)。[20] H. Liu,J。Weng,胶质瘤中细胞周期蛋白依赖性激酶2(CDK2)的全面生物信息学分析,Gene,(2022)146325。[21] H. Liu,T。Tang,Pan-Cancer的库糖胞化和铜代谢相关的基因集,肿瘤学的边界,12(2022)952290。[22] H. Liu,Y。Li,Cornichon家族AMPA受体辅助蛋白4(CNIH4)在头部和颈部鳞状细胞癌中的潜在作用,癌症生物标志物:疾病标志物A部分(2022)。[23] H. Liu,J.P。Dilger,J。Lin,pan-Cancer-Biodorminicals-Informinical-Informicals Trpm7的文献综述,Pharmacol Ther(2022)108302。[24] H. Liu,cuproptosis Gene Set的Pan-Canter概况,《美国癌症研究杂志》,第12期(2022)4074-4081。[25] Y. Liu,H。Liu,氨基酰基TRNA合成酶复合物的临床能力相互作用多功能蛋白1(AIMP1),用于头颈鳞状细胞癌,癌症生物标志物:疾病标志物A节A节(20222)。[26] Y. Li,H。Liu,Y。Han,在头部和颈部鳞状细胞癌中,Cornichon家族AMPA受体辅助蛋白4(CNIH4)的潜在作用,研究方形(2021)。 '

人类癌症中功能性A-to-I RNA编辑事件的综合研究
摘要 腺苷到肌苷 (A-to-I) RNA 编辑占人类所有 RNA 编辑事件的近 90%,据报道它与多种癌症的致瘤性有关。然而,对癌症中功能性 A-to-I RNA 编辑事件的全面图谱仍然不足。为了填补这一空白,我们系统而深入地分析了来自 Cancer Genome Atlas 的 33 种癌症样本中 A-to-I RNA 编辑事件的多种致瘤机制。对于 1,500,000 个量化的 RNA 编辑事件中的个别候选事件,我们进行了不同类型的下游功能注释。最终,我们确定了 24,236 个潜在功能的 A-to-I RNA 编辑事件,包括 APOL1、IGFBP3、GRIA2、BLCAP 和 miR-589-3p 中的案例。这些事件可能在肿瘤发生过程中发挥关键作用,因为它们具有与肿瘤相关的编辑频率,或可能对改变的表达谱、蛋白质功能、剪接模式和肿瘤基因的 microRNA 调控产生影响。我们的功能性 A-to-I RNA 编辑事件 ( https://ccsm.uth.edu/CAeditome/ ) 将有助于从 A-to-I RNA 编辑角度更好地了解癌症病理学。

以细胞线粒体自噬为目标作为治疗人类癌症的策略
线粒体自噬是细胞选择性清除功能失调的线粒体的过程,控制着线粒体的数量和质量。线粒体自噬失调可能导致受损线粒体的积累,在肿瘤的发生和发展中起着重要作用。线粒体自噬包括由PINK1 / Parkin介导的泛素依赖性途径和由线粒体自噬受体(包括NIX,BNIP3和FUNDC1)介导的非泛素依赖性途径。细胞线粒体自噬广泛参与多种细胞过程,包括代谢重编程,抗肿瘤免疫,铁死亡以及肿瘤细胞与肿瘤微环境之间的相互作用。并且细胞线粒体自噬还调节肿瘤的增殖和转移,干细胞,化学抗性,对靶向治疗和放射治疗的抵抗力。在这篇综述中,我们总结了线粒体自噬的潜在分子机制,并讨论了线粒体自噬在不同肿瘤环境中的复杂作用,表明它是线粒体自噬相关抗肿瘤治疗的一个有希望的靶点。

人类癌症遗传学计划
该计划与临床界密切合作,以促进基因诊断和研究方面的合作,并促进培训和教育。2023 年,家族性癌症咨询中心共接待了 1,145 名患者(579 例新病例和 566 名患者接受结果),HCGP 进行了 2,128 次基因诊断并进行了 2,102 项细胞遗传学研究。在研究方面,该计划的成员参与了涉及我们国家卫生系统 30 多家医院网络的合作研究,不仅开展合作研究,还促进临床转化。该计划还为来自不同国家和国际研究中心的专业人士和学生提供加入的机会,无论是作为访问者还是进行短期停留 1 至 3 个月的培训访问。2023 年接待了 3 名国内访客和 7 名国际访客,以及来自不同西班牙医院的 4 名住院医师。在教育方面,5 名本科生、2 名硕士生和 16 名博士生参与了他们的研究项目。

人类癌细胞中肉瘤外DNA物种的协调遗传
超颞神经。b。颤振设备的3D打印部分。c。振动装置的组装。RCA电缆通过B5并提供振动电动机。d。调查结果有关佩戴纤维状刺激装置的经验。e。实验平台。参与者坐在椅子上,并用占主导地位的手握住按钮。f。本研究中使用的N-BACK任务的例证。对于0-back任务,要求参与者看到0。在2退的任务中,当参与者认为当前呈现的数字与两次试验之前的数字相同时,他们被指示做出回应。g。n在基线会话中的范围从1到8。VN和假会话包括八个块,每个块都有一个用于0折叠任务的块,一个用于1折叠任务,而两个块则用于2、3和4-back任务。