XiaoMi-AI文件搜索系统
World File Search System羔羊

断奶羔羊
这项随机对照疫苗实地试验的目的是确定一种商用呼吸道疫苗 Ovipast ™ Plus 的效果,该疫苗在断奶时(约 8 周龄)施用于饲养场羔羊,以改善动物健康、生长性能和胴体性状。羔羊断奶、称重,并根据性别和母亲之前的 Ovipast ™ Plus 疫苗接种情况分组,因为这是正在进行的试验的第 2 阶段(Gardner 等人,2023 年)。接种疫苗的母羊所生羔羊接种了 Ovipast ™ Plus 疫苗,并在 3 – 5 周后再次接种疫苗。未接种疫苗的母羊所生羔羊未接种疫苗。在试验的生长和育肥阶段,接种疫苗不会降低肺炎治疗率、粗死亡率或肺炎特定死亡率,也不会提高生长率。与未接种疫苗的羔羊相比,接种疫苗的羔羊胴体脂肪覆盖率较低(P < 0.001),产出 1 级胴体的几率增加 1.33 倍(P = 0.01)。接种疫苗降低了胴体脂肪覆盖率并提高了产量等级,但对疾病率或生长性能没有有益影响,这表明在断奶后接种疫苗的羔羊的经济效益有限,这些羔羊是由在妊娠期间接种 Ovipast ™ Plus 的母羊所生。


初级市场羔羊师
Division Champion & Reserve Champions ............................................................................................................................................................ Ban ner Grand Champion Market Lamb .................................................Banner & Buckle by NWSS Reserve Grand Champion Market Lamb ......................................NWSS国家西部股票节目 Banner&Buckle展示了Charles E. Kirk Memorial Belt Buckle,向大冠军市场羔羊的获胜者。 预备役冠军也将获得皮带扣。Banner&Buckle展示了Charles E. Kirk Memorial Belt Buckle,向大冠军市场羔羊的获胜者。预备役冠军也将获得皮带扣。

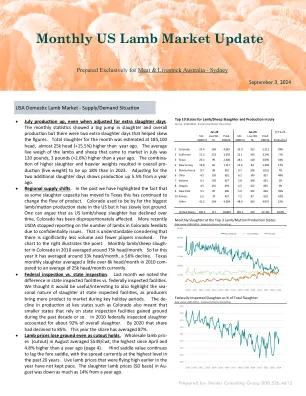
每月美国羔羊市场更新
•7月份的生产,即使调整了额外的屠杀日。每月的staɵsɵcs在屠宰和整体生产中表现出了很大的跳跃,但额外的屠杀日有助于歪曲这些数字。本月的总屠杀率为185,100头,比一年前高25,000头(+15.5%)。7月份上市的羔羊和绵羊的平均活体重为120磅,比去年同期高3磅(+2.6%)。高屠杀和重量更重的组合导致总体疾病(活体重)的增长比2023年增长18%。Adgusɵng在两个添加的屠杀日期表明,生产率比去年同期增长了5.5%。区域供应shiōs。过去,我们强调了一个事实,即随着某些屠宰能力已搬到德克萨斯州,这已经引起了改变产品的影响。科罗拉多州曾经是迄今为止美国最大的羔羊/穆恩(Muʃon)的生产州,但它逐渐失去了地面。可以说,随着我们的羔羊/绵羊的屠杀,科罗拉多州的屠宰已经下降了。最近,美国农业部因征服性问题而停止了科罗拉多饲养场中羔羊的数量。可以理解的是,考虑到数量较小,参与者较少。右侧的图表说明了点。2010年在科罗拉多州在科罗拉多州的每月羔羊/绵羊Slaughter slaughter平均每月约为75,000。今年到目前为止,平均每月约为33K,下降了56%。德克萨斯州月度屠杀平均每月超过8,000,目前平均每月为25,000。联邦Inspecɵon与州Inspecɵon。上个月,我们注意到国家检查的区别与联邦政府检查。我们认为,随着生产商在关键的假期期间将更多的产品推向市场,这也可以强调州检查的屠杀的海洋性质是有用的。在科罗拉多州等关键州的生产中的销售也意味着依赖于过去十年左右的较小的州的较小州在占领的情况下获得了地面。在2010年,联邦政府检查的屠宰约占整体屠宰的92%。到2020年,该份额已下跌至85%。今年的份额平均为87%。•羔羊价格即使裁切持有,也会损失地面。8月的批发羊肉定价(切口)平均为549美元/cwt,这是4月以来最高的,比一年前高4.8%(第4页)。Hind Maddle的价值会落后于鞍座,过去20年中的差异目前处于最高水平。在今年早些时候高涨的活羔羊价格并没有保持步伐。Au -Gust的屠宰羔羊价格(SD基础)比去年同期下降了14%。


测试目录-Enniscorthy
祖先信息1)替换(¼3.44)±如果农民希望公羊两者都繁殖羔羊以杀死屠宰和替换母羊羔羊,那么他们应该根据该指数选择RAM。该指数的大部分是羔羊生存能力的组合,屠杀的日子,编号羔羊出生和女儿牛奶。索引旁边的¼值表示从该RAM饲养的每个羔羊的利润差异。此¼值是跨品值,因此可以将不同的品种相互比较。2)末端(¼2.86)±如果农民希望只有公羊只繁殖羔羊以杀死屠宰,则应根据该指数选择RAM。该指数的大部分是羔羊生存能力和屠杀天数的组合。索引旁边的¼值表示从该RAM饲养的每个羔羊的利润差异。跨品值的此¼ydoxh lv an the ydoxh lv,因此可以将不同的品种彼此进行比较。3)羔羊的生存能力±这基于羔羊的易感性和羊羔的存活率对动物进行排名。该指数上的每个正百分点都是一个额外的羔羊,可以存活40天,反之亦然。4)屠杀天数±这对动物的遗传能力进行了排名。与该性状相关的指数表示与同一品种的其他动物相比,该公羊后代达到所需的屠宰体重所需的预测天数差异。5)编号对动物进行了对动物的排名。6)女儿牛奶±这与同一品种的其他动物相比,根据其女儿的预测挤奶表现对动物进行排名。

...
由于Worlton [1]使用羔羊波来非破坏性测试板,因此对NDT中的板波的应用引起了极大的兴趣。羔羊波可在局部化中使用。详细的NDT应用程序。可以通过在适当的频率厚度产品中选择最合适的模式来优化给定缺陷的可检测性。同样,由于它们本质上是二维,因此羔羊波的衰减速度低于3维散装波。,因此可以在相当长的距离内传播。因此。lat-rib波可在远程NDT应用中使用。可以将大型板状结构进行粗略检查。但是。始终存在多个传播羔羊波,并且速度分散通常很明显。因此,耀斑对施加的兴奋剂的响应的时间历史只能用于大约测量羔羊波幅度和ve弹性。因为响应信号的形状将在沿板表面的不同位置处有不同的位置。也是如此。如果羔羊模式的组速度相似,则需要在可以在时域中解析之前,需要坚持的繁殖距离。

腹泻(搜寻)小反刍动物
球虫病是一种原生动物寄生虫病,是羔羊和儿童腹泻的常见原因。它也可能导致亚临床生产损失。羔羊和孩子在1至4个月大时对这个问题最感染,尽管可能受到年轻动物的影响。羔羊在生命的头几周对疾病具有抗性。在此期间暴露于原生动物中,赋予了对后来感染的免疫力和抵抗力。临床疾病在断奶,饲料变化或运输的压力后很常见。羔羊和孩子的腹泻通常不是血腥的,但可能含有血液或粘液,非常水。对受影响动物的治疗包括支持性护理和coccidiostats的助教。一组动物应在爆发期间接受治疗。预防涉及改善卫生和使用Coccidiostats。

母体剥夺和牛奶的替代影响正在发育中的羔羊脑中灰色和白质的完整性
侵略性,以及一般的刻板印象行为,例如非果实哺乳,头部打击或自我伤害(Latham&Mason,2008)。母亲剥夺的这些影响可能是由于缺乏社会模型(即母亲(Fleming等,2002))以及缺乏母乳作为生物活性因素的来源(Bernstein&Hinde,2016年)而同时导致的。然而,挑战不是要隔离一个因素(母亲或牛奶)的影响,而是考虑“同样重要的母亲和非母性变量之间的动态相互作用”(Tang等,2014)。在绵羊(ovis aries)中,一种早熟的物种,很难将母体剥夺的影响与配方奶粉喂养的影响分解。每个都影响婴儿的发育。母乳和她的羔羊之间的互惠母亲 - 在生命的前12小时发生。它主要基于气味线索,其特点是对每个伴侣的个人认识和母亲和她自己的婴儿之间的护理排他性(Nowak等,1997,2011; Nowak&Poindron,2006)。在这种情况下,母亲对于羔羊的发育至关重要,这并不奇怪。但是,无母亲的饲养通常用于常规乳制品耕作,或者在其他情况下,如果母亲是非母亲,则羔羊过多或患有乳腺炎。尽管在绵羊种植方面具有普遍的做法,但涉及母亲剥夺,牛奶替代或早期断奶的早期饲养条件的影响会影响广泛的功能和行为。这些研究证明了母亲的剥夺和结束 -出生时,由于从母亲到羔羊的被动免疫转移,初乳对羔羊的生存很重要(Hernández-Castellano等,2015; Khan&Ahmad,1997; Nowak&Poindron,2006年)。与富含母体和商业奶的混合物相比,用商业牛奶替代品喂食的羔羊的免疫反应改变了(Sevi等,1999)。然而,在出生后几天被剥夺了母亲,而不是出生时,羔羊可以进入初乳,并大大降低了对免疫反应的影响,特别是如果随后用母羊的牛奶喂养(Napolitano,2003; Napolitano等,1995年)。在生命的头几周里,母亲是一个社会示威者的关键作用,影响了喂养的建立(Black-Rubio等,2007; Saint-Dizier等,2007; Thorhallsdottir等,1990)和双胞胎之间的社会偏好(Ligout&Porter,2004)或Appeasepe fy(Ligout&Porter,2004)或appeaseme ty Al caregiv al Al al Al an。 )。从长远来看,母亲的缺失对男性羔羊的性行为表达产生了负面影响(Damián等,2015,2018)。情感反应性在社会隔离环境中通过皮质醇血浆水平和行为反应评估,也受到母性剥夺的影响(Napoli- Tano,2003; Napolitano等,2002; Sevi等,1999)。另外,还报道了内分泌不平衡的性行为行为(Damián等,2015,2018),婴儿依恋(Gaudin等,2018)或营养(Berry等,2016)。