XiaoMi-AI文件搜索系统
World File Search System脑前

枳果梗多糖调节谷氨酸代谢和紧密连接蛋白表达改善酒精 ...
摘 要 : 目的:本研究旨在明确枳椇果梗多糖( HDPs )对酒精暴露所致的小鼠神经行为异常的改善效果,并探究谷 氨酸代谢和紧密连接蛋白表达在其中的作用。方法:雄性 C57BL/6 小鼠按 114 μL/20 g 剂量连续酒精灌胃 14 d ,建 立酒精暴露模型,同时设置干预组进行 HDPs 干预( 114 μL/20 g 酒精 +100 mg/kg HDPs )。应用行为学实验(旷场 实验、高架十字迷宫实验)评估神经行为学变化,采用气相色谱法测定小鼠血液中乙醇浓度, γ -H2AX 荧光检测小 鼠脑海马组织 DNA 损伤,免疫组化分析检测小鼠脑组织中紧密连接蛋白 Claudin-1 和 ZO-1 的表达,并通过超高 效液相色谱 - 四级杆飞行时间质谱法( UPLC-Q-TOF-MS )代谢组学技术对小鼠脑组织代谢物进行分析。结果: HDPs 可有效降低酒精暴露小鼠血液乙醇浓度,由 4.69±0.29 g/L 降至 1.64±0.104 g/L ;改善酒精暴露所致的小鼠神 经行为异常,旷场实验中,与酒精组相比, HDPs 干预组总路程显着提升至 27340±3304 cm ( P <0.05 ),平均速度 显着提升至 67.4±13.4 cm/s ( P <0.05 ),不动时间缩短 29% ( P <0.05 );高架十字迷宫实验中,与酒精组相比, HDPs 干预组闭臂停留时间显着减少至 195.6±10.3 s ( P <0.05 ),开放臂进入次数显着增加 26% ( P <0.05 ));还 可降低酒精诱导的脑组织氧化应激与 DNA 损伤水平, ROS 、 MDA 分别降低 5.4% 、 29.5% ( P <0.05 ), T-AOC 提 高 10.9% ,上调脑海马组织中 Claudin-1 ( 2.2 倍)和 ZO-1 ( 0.1 倍)蛋白的表达;并调节脑组织谷氨酸代谢通路, 提高甘氨酸( 19.7% )、谷光甘肽( 25% )、琥珀酸( 22.6% )等代谢物水平。结论: HDPs 可有效改善酒精对小鼠 神经行为的影响,其机制或可能通过抗氧化、保护紧密连接蛋白和调节谷氨酸代谢通路发挥作用,研究结果可为 扩展枳椇资源在食品领域中的应用提供理论依据。


学习和先天威胁规避的全脑投射和前额叶神经元类别的差异编码
要了解大脑如何产生行为,我们必须阐明神经元连接与功能之间的关系。内侧前额皮质 (mPFC) 对决策和情绪等复杂功能至关重要。mPFC 投射神经元广泛侧支,但 mPFC 神经元活动与全脑连接之间的关系尚不清楚。我们进行了全脑连接映射和光纤光度测定,以更好地了解控制雄性和雌性小鼠威胁回避的 mPFC 回路。使用组织透明化和光片荧光显微镜 (LSFM),我们绘制了投射到伏隔核 (NAc)、腹侧被盖区 (VTA) 或对侧 mPFC (cmPFC) 的 mPFC 神经元群的全脑轴突侧支。我们提出了 DeepTraCE(基于深度学习的追踪与综合增强)来量化透明组织图像中批量标记的轴突投射,以及 DeepCOUNT(基于深度学习的通过 3D U-net 像素标记进行物体计数)来量化细胞体。使用 DeepTraCE 生成的解剖图与已知的轴突投射模式对齐,并揭示了区域内类别特定的地形投射。使用 TRAP2 小鼠和 DeepCOUNT,我们分析了威胁回避背后的全脑功能连接。PL 是与 PL-cPL、PL-NAc 和 PL-VTA 目标位点子集具有功能连接的最高度连接的节点。使用光纤光度法,我们发现在威胁回避过程中,cmPFC 和 NAc 投射器编码条件刺激,但仅在需要采取行动避免威胁时才会编码。mPFC-VTA 神经元编码学习到的但不编码先天的回避行为。总之,我们的研究结果为定量全脑分析提供了新的和优化的方法,并表明解剖学定义的 mPFC 神经元类别在避免威胁方面具有特殊的作用。

rab6介导的突触囊泡前体的极化转运对于建立神经元极性和脑形成至关重要
神经元是由单个轴突和多个树突组成的高度极化细胞。轴突 - 树突极性对于正确的组织形成和脑功能至关重要。细胞内蛋白转运在神经元极性的建立中起重要作用。但是,极化运输的调节机制尚不清楚。在这里,我们表明Rab6是一种针对细胞内囊泡传统调节的小GTPase,在神经元极化和脑发育中起着关键作用。中枢神经系统特异性RAB6A/B双敲除(RAB6 DKO)两性的小鼠均表现出新皮质和小脑的严重发育不良。在Rab6 DKO新皮层中,神经元的轴突延伸受损会导致中间区发育不全。在体外,从性别中培养的神经元中Rab6a和Rab6b的缺失会导致与高尔基体相邻的突触囊泡前体(SVP)的异常积累,从而导致轴突延伸中的缺陷和Axon -Axon -dendrite Polarity的丧失。此外,Rab6 DKO会导致神经元中溶酶体的显着膨胀。总体而言,我们的结果表明,RAB6介导的SVP的极化转运对于神经元极化和随后的脑形成至关重要。

小鼠终纹外侧和内侧前背床核的全脑输入映射
终纹床核 (BNST) 的前部调节恐惧和压力反应。前背 BNST (adBNST) 在解剖学上可进一步细分为外侧和内侧部分。尽管已经研究了 BNST 亚区的输出投影,但对这些亚区的局部和全局输入连接仍然知之甚少。为了进一步了解以 BNST 为中心的电路操作,我们应用了新的病毒遗传追踪和功能电路映射来确定小鼠 adBNST 外侧和内侧亚区的详细突触电路输入。在 adBNST 亚区注射了单突触犬腺病毒 2 型 (CAV2) 和狂犬病毒逆行示踪剂。杏仁核复合体、下丘脑和海马结构占 adBNST 总体输入的大部分。然而,外侧和内侧 adBNST 亚区具有不同的长距离皮质和边缘大脑输入模式。外侧 adBNST 具有更多来自前额叶(前边缘、下边缘、扣带回)和岛叶皮质、前丘脑和外嗅皮层/外嗅皮层的输入连接。相比之下,内侧 adBNST 接收来自内侧杏仁核、外侧隔膜、下丘脑核和腹侧下托的偏向输入。我们使用 ChR2 辅助电路映射确认了从杏仁海马区和基底外侧杏仁核到 adBNST 的长距离功能输入。选定的新型 BNST 输入还通过来自艾伦研究所小鼠脑连接图谱的 AAV 轴突追踪数据进行了验证。总之,这些结果提供了外侧和内侧 adBNST 亚区差异传入输入的全面图谱,并为 BNST 电路对压力和焦虑相关行为的功能操作提供了新的见解。
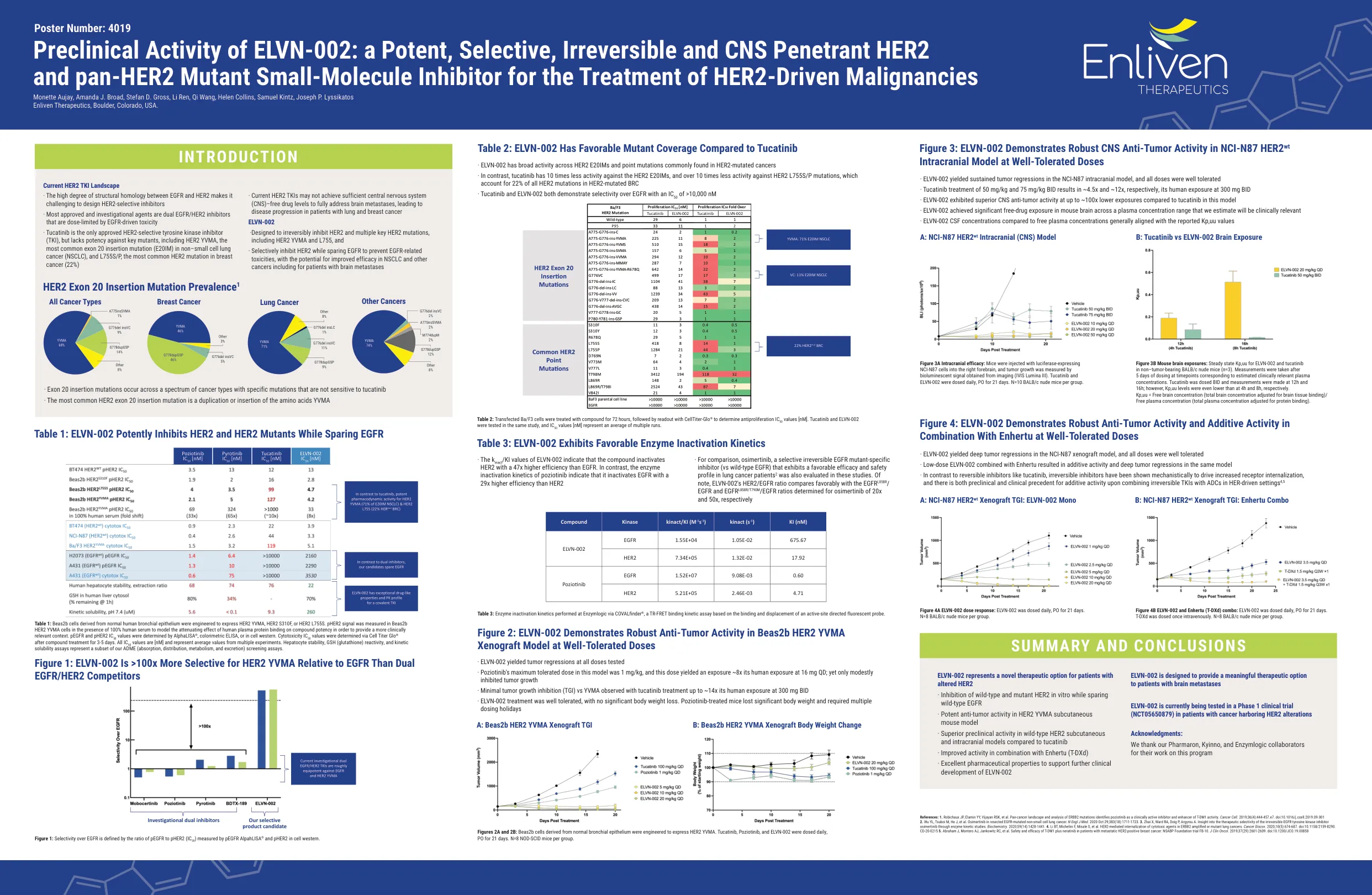
ELVN-002的临床前活动ELVN-002的临床前活动
表1:衍生自正常人支气管上皮的BEAS2B细胞被设计为表达HER2 YVMA,HER2 S310F或HER2 L755。PHER2信号,以建模人血浆蛋白结合对复合效力的衰减作用,以提供更临床相关的环境。PEGFR和PHER2 IC 50值通过alphalisa®,比色ELISA或细胞西部确定。细胞毒性IC 50值是通过复合处理后通过细胞滴度GLO®确定的3-5天。所有IC 50值都是[NM],代表来自多个实验的平均值。肝细胞稳定性,GSH(谷胱甘肽)反应性和动力溶解度测定代表了我们ADME(吸收,分布,代谢和排泄)筛选的子集。

植入式柔性神经微电极最新进展
∗ 基金项目 : 科技创新 2030“ 脑科学与类脑研究 ” 重大项目 (2022ZD0208601), 国家自然科学基金 (62076250,62204204), 陕西



公告前海军航空站...
修复咨询委员会会议日期:2024 年 10 月 2 日,星期三时间:下午 6:00 - 晚上 8:00 地点:仅通过互联网进行虚拟访问:https://tinyurl.com/NASBOCT24RAB 会议 ID:254 668 103 758 密码(区分大小写):VAvfxr 电话:+1 877-286-5733 电话会议代码:629 543 152# 海军设施工程系统司令部基地调整和关闭计划管理办公室 (NAVFAC BRAC PMO) 宣布召开前海军航空站布伦瑞克 (NASB) 修复咨询委员会 (RAB) 会议。会议将仅以虚拟形式举行,可通过上面提供的 Teams 会议链接访问。海军将提供正在进行的活动的更新或状态审查,包括全氟和多氟烷基物质相关活动、长期监测、沉积物和地下水修复相关活动、土地使用控制、财产转让和社区外展工作。会议材料将在线提供,网址为 https://www.bracpmo.navy.mil/BRAC-Bases/Northeast/Former-Naval-Air-Station- Brunswick/Meeting-Material/ 。如有疑问或需要更多信息,请联系 BRAC PMO 东部环境协调员:W. Rachelle Knight 女士,BRAC 环境协调员,BRAC PMO 东部,4911 South Broad Street,Building 679,Philadelphia,PA 19112,电话 (215) 897-4916 或电子邮件 wynette.r.knight.civ@us.navy.mil。