XiaoMi-AI文件搜索系统
World File Search System酸酸

在全反触手酸酸诱导的神经元分化
摘要:人神经母细胞瘤细胞系SH-SY5Y和IMR-32可以通过用全反替酸(ATRA)处理分化为神经元样的表型。分化后,这些细胞系被广泛用作体外模型来研究神经元细胞生物学的各个方面。然而,在ATRA诱导的分化中,SH-SY5Y和IMR-32细胞的蛋白质组和磷酸蛋白质组的时间和定量分析受到限制。在这里,我们在ATRA诱导的分化过程中,在多个时间点对SH-SY5Y和IMR-32细胞的蛋白质组和磷酸蛋白质组进行了相对定量。与随后的基因本体分析的蛋白质和磷酸肽的相对定量表明,包括细胞骨架组织,细胞分裂,伴侣伴侣功能和蛋白质折叠以及单碳代谢在内的几种生物学过程与两种细胞系中ATRA诱导的分化都相关。此外,激酶 - 基底富集分析预测了分化过程中几种激酶的活性改变。其中,CDK5表现出增加的活性,而CDK2的活性降低。提供的数据是研究在ATRA诱导的分化过程中SH-SY5Y和IMR-32细胞中时间蛋白和磷蛋白丰度变化的宝贵资源。

通过调节假ex胶剪接模式来调节PCCA基因表达,以挽救丙酸酸血症中的酶活性
伪exon是非功能性内含子序列,可以通过深内核序列变化激活。激活中的伪exon包含在mRNA中,并干扰了正常的基因表达。PCCA C.1285-1416A> g变化激活伪exon,并通过在PCCA和PCCB中编码的丙酰基-COA羧化酶酶的指示引起严重的代谢性毒性酸血症。我们详细介绍了这种致病性伪exon活化事件,并确定HNRNP A1对于正常代表很重要。PCCA C.1285-1416A> g变化破坏了HNRNP A1结合剪接消音器,并同时创建剪接增强器。我们证明,通过剪接切换的反义寡核苷酸阻止这种调节区域可恢复正常的剪接,并挽救患者纤维细胞中的酶活性,并在由CRISPR基因创建的细胞模型中恢复了酶活性。有趣的是,PCCA伪exon具有上调基因表达的未插入潜力,因为健康组织显示出相对较高的纳入水平。通过阻止未激活的野生型假exon的包含,我们可以同时增加PCCA和PCCB蛋白水平,从而增加了异二次运动酶的活性。令人惊讶的是,我们可以从具有PCCA错义变体的患者纤维细胞中的残留水平中吸收酶活性,而且还可以从具有PCCB错过变体的患者中进行酶活性。这是丙酸血症的潜在治疗策略。
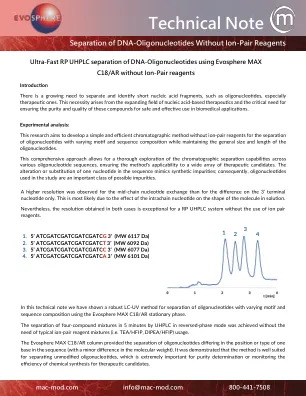
使用IVosphere Max C18-AR不含离子对试剂的超快速RP UHPLC分离DNA-寡核苷酸
总而言之,该研究涉及对能够分离和鉴定短核酸片段(尤其是治疗性寡核苷酸)的高级色谱方法的紧迫需求。通过使用C18AR色谱柱进行系统评估,具有不同基序和序列组成的寡核苷酸,以及模仿序列杂质的掺入,可以增强可用的分析工具,以确保基于核酸酸的治疗剂的质量和安全性。

PFAS来源调查和识别计划...
来源调查和识别工作的目的是识别和描绘促进和多氟烷基物质(PFAS)的贡献领域,即见于全氟辛烷磺酸(PFOS)和全氟辛酸酸(PFOA)(PFOA)(PFOA)(PFOA),以最佳的危害者的发展和其他适用的污染物,并为开发和实施做好了正式的采取行动。目标是满足PFA的水质值(WQV),以排出该州的水域。

S445-100肉桂streusel Topping
成分糖,富含粉的粉(小麦粉,烟酸,降铁,硫胺素单硝酸盐,核黄素,叶酸),谷物碎屑(浓粉(富集面粉[小麦粉,烟酸,降低铁,硫胺素,硫胺素单硝酸盐,单硝酸盐),单硝酸盐,单硝酸盐,核酸果酸,核酸酸酯,叶酸] ,黄油(巴氏杀菌奶油,天然风味),大豆油,水,甘蔗糖蜜,肉桂。

N390-300 RainbowNerds®
成分玉米糖浆,糖,葡萄糖含有2%或更少的苹果酸(DL-麦酸酸),口香糖阿拉伯语,天然风味,康巴纳蜡,添加了颜色,黄色5,红色40湖,50湖,黄色5湖,蓝色湖泊,蓝色2湖,蓝色湖泊,蓝色1,carmine 6,颜色1,蓝色1,蓝色1,蓝色1,蓝色1,蓝麦汁浓缩。

brilinta-rododuct-monograph-en.pdf
急性冠状动脉综合征Brilinta(Ticagrelor)与低剂量乙酰基酸酸(ASA:75-150 mg)共同管理,以预防急性冠状动脉综合征(ACS)患者的动脉粥样硬化事件(请参阅14个临床试验)。与低剂量的乙酰水杨酸(ASA:75-150毫克)共同管理的Brilinta(ticagrelor)(ticagrelor)的历史,表明次要预防次要预防在有症状的病史的患者(MI)发生的一年及其一年的患者(至少是MI)的次数预防(至少是MI)。 (请参阅14次临床试验)。冠状动脉疾病,2型糖尿病和经皮冠状动脉介入疗法的历史,与低剂量乙酰基酸酸(ASA:75-150毫克)共同采用,表明与第一次心肌肌肉症患者(Coronary Artter)患者降低了Coronary ARTORE ARARARY ARTORE ARARARY ARIDARE ARARICTIAL ARARARY ARIDERARE ARARARY ARARINARY ARIDERARY ARARICTIAL ARARICTIAL ARARICTIAL ARARICTIAL ARTERARY ARTORE CORONARY ARTORE cORONARARY ARIDER, Mellitus(DM)和经皮冠状动脉干预史(PCI)的史,他们也有发展动脉粥样硬化事件的高风险(请参阅4剂剂量和管理,7项警告和预防措施以及14项临床试验)。

生物生产乳酸
乳酸酸已经出现在商业现场,是一种多功能的多羟基酸,在食品,药品,药物,化妆品和化学工业中都有许多合理的应用。这种高增值的生物产品最近作为生物活性化合物越来越受到关注,为合成新型潜在的生物相容性和可生物降解的药物脱脂车提供了出色的化学平台。组织工程和纳米医学的最新进展也强调了该有机酸作为关键生物功能化剂的重要性。因此,乳酸酸的商业相关性不断增长,促使其生物技术生产的新型系统既可持续又有效。本评论探讨了与乳酸生物生产有关的最新进展和研究,无论是通过微生物还是酶促方法,突出了增强生物生产的关键生物处理条件。还列出了当前微生物细胞工厂的详细概述以及乳酸生产的下游加工方法。此外,还讨论了该多羟基酸的潜在前景和当前应用,重点是乳酸离子酸作为新型药物,生物瘤,纳米颗粒和生物聚合物系统开发的关键平台的作用。©2013 Elsevier Inc.保留所有权利。

人类心力衰竭中的类花生素:血浆环氧树叶和二羟基乙酸的试验研究
心力衰竭(HF)是心血管发病率和死亡率的主要原因,随着患病率的增加,全球医疗系统面临HF大流行[1-3]。尽管现代药物疗法,包括血管紧张素受体 - 抑制剂抑制剂(ARNI)和 - 葡萄糖糖共转移蛋白-2抑制剂(SGLT2I),但患者的预后,尤其是患有晚期HF的人的预后仍然很差[4]。eicosanoids先前在基本的心血管和肾脏研究中进行了研究。这些细胞色素P-450(CYP) - 脱发 - 烯烃的代谢产物(AA),尤其是环氧酸 - 辛酸 - 辛酸 - 辛酸酸(EET),重要的是,重要的是,通过其vasodilital and natriuration and natriuration和Natriurater效应,有助于调节car骨和肾脏系统。此外,在临床前研究中,它们发挥了器官保护作用[5-7]。在生理条件下,EET由内皮细胞(作为内皮衍生的超极化因子 - EDHF)和表现出自分泌和旁分泌作用而产生。eets被可溶性环氧水解酶(SEH)转化为生物学上的活性较低的二羟基乙酸酸酯(DHETS)[5,8],并主要排出