XiaoMi-AI文件搜索系统
World File Search System黄籽

从富营养化池塘中分离的絮凝黄杆菌(Luteibacter flocculans sp. nov.)以及 LuteibacterPhage vB_LflM-Pluto 的分离和表征
Luteibacter 属是 Rhodanobacteraceae 科的一部分,属于变形菌门的 γ 亚纲。该科包含 17 个属,分别是 Aerosticca、Ahniella、Aquimonas、Chiayiivirga、Denitratimonas、Dokdonella、Dyella、Frateuria、Fulvimonas、Luteibacter、Oleiagrimonas、Pinirhizobacter、Pseudofulvimonas、Rehaibacterium、Rhodanobacter、Rudaea 和 Tahibacter,其中两个属尚未有效发表(Denitratimonas 和 Pinirhizobacter)[1]。Luteibacter 属由 Johansen 等人 [2] 基于 Luteibacter rhizovicinus DSM 16549 T 种建立。该属目前包含 5 个种,其中 3 个已有效发表:L. rhizovicinus DSM 16549 T [ 2 ]、L. yeojuensis DSM 17673 T [ 3 , 4 ]、L. anthropi CCUG 25036 T [ 4 ],以及 L. jiangsuensis [ 5 ] 和 L. pinisoli [ 6 ]。Luteibacter 属的成员分离自各种环境,例如根际土壤 [ 2 , 6 ]、温室土壤 [ 3 ] 和人体血液 [ 4 ]。它们被描述为具有运动能力的、需氧的革兰氏阴性菌,呈杆状,呈黄色。此外,它们是过氧化氢酶和氧化酶阳性和脲酶阴性的。迄今为止,Luteibacter 或甚至是 Rhodanobacterceae 相关噬菌体都是未知的。噬菌体或细菌噬菌体是感染细菌的病毒。虽然温和噬菌体可以整合到细菌基因组中,但溶菌噬菌体在感染后直接开始繁殖。温和噬菌体会将其整合的基因组与宿主基因组一起复制,从而产生原噬菌体和溶原性细菌。通过添加其遗传物质,原噬菌体可以提供新的能力,保护宿主免受相关和不相关病毒的感染 [ 7 ]。在之前的研究中,我们从位于德国哥廷根的一个富营养化池塘中分离出一种环境 Luteibacter sp. nov. 菌株。分离 Luteibacter 菌株作为预期模型菌株,以研究与细菌感染相关的局部病毒多样性。

鹦鹉鲜艳色彩变化的分子机制
结果:通过结合光谱、色谱和质谱分析不同物种的羽毛,我们发现了鹦鹉从黄色到红色颜色变化的共同化学基础。我们发现鹦鹉黄霉素“端基”的氧化状态在颜色变化中起着关键作用,颜色从黄色到红色的调整与鹦鹉黄霉素分子中羧基与醛基端基的比例相关;红色羽毛含有大量的醛基鹦鹉黄霉素,而黄色和绿色羽毛的羧基鹦鹉黄霉素含量较高。为了探索这些颜色差异的遗传基础,我们研究了暗色鹦鹉,它在野生种群中有两种:黄色和红色。遗传图谱确定了一个与颜色变化相关的基因组区域,其中包含 ALDH3A2 基因下游非编码区域中的候选点突变,该基因编码一种催化氧化的酶

Minutes - 立法会
立法会 立法会 立法会文件编号 CB(2)1150/10-11 (本会议记录已由政府当局查阅) 编号:CB2/PL/SE 保安事务委员会 2011 年 1 月 17 日星期一上午 8 时 45 分在立法会大楼会议厅举行的特别会议记录 成员: 涂谨申议员 (主席) 出席 刘江华议员, JP (副主席) 何俊仁议员 吴霭仪议员 张文光议员 黄容根议员, SBS, JP 刘慧卿议员, JP 霍震霆议员, GBS, JP 石礼谦议员, SBS, JP 余若薇议员, SC, JP 林大辉议员BBS, JP 叶国谦议员, GBS, JP 潘佩秋议员 梁国雄议员 黄毓民议员 委员: 李卓人议员 出席 黄国兴议员, MH 议员: 黄裕康议员, GBS 缺席 梁君源议员, GBS, JP 詹议员佩忠议员何秀兰议员陈克勤议员黄国健议员, BBS 谢伟俊议员

HAL 的移动多平台图集
在本文中,我们建议通过不同的在线平台(Facebook、Twitter 和 YouTube)绘制在线社会运动,更准确地说是黄背心运动。我们的目标是双重的:一方面建立黄背心在 Facebook 群组中的活动地图集,另一方面捍卫一种尽可能混合在不同数字平台上收集的痕迹的方法论。黄背心 Facebook 群组将成为我们对该运动的特权观察点。如果对 Facebook 上发布的帖子内容的检查,再加上对每个群体附带的大量元数据的分析,使我们能够界定运动需求空间的轮廓及其聚合动态,我们利用这些出版物中传播的链接来使关于该运动的观点多样化。因此,我们根据在微博平台上转发这个或那个 URL 的人计算出 Twitter 的政治倾向。黄背心在网络上引用外部资源的方式(无论是网站还是 YouTube 频道)也揭示了他们与媒体的关系。因此,我们依靠数字公共空间中主要媒体的现有特征来理解它们与主流、激进媒体(无论是右翼还是左翼)或反信息空间的关系。分析

硬件黑客 - kea.nu
名称:黄,安德鲁,作者。标题:硬件黑客:制造和破解硬件的冒险 / 安德鲁“Bunnie”黄。说明:第 1 版。| 旧金山:No Starch Press,Inc.,[2017] 标识符:LCCN 2016038846(印刷版)| LCCN 2016049285(电子书)| ISBN 9781593277581(平装本)| ISBN 159327758X(平装本)| ISBN 9781593278137(epub)| ISBN 1593278136(epub)| ISBN 9781593278144(mobi)| ISBN 1593278144(mobi)主题:LCSH:电子设备和电器 - 设计和构造。|电子设备和器具——技术创新。| 计算机输入输出设备——设计和构造。| 逆向工程。| 电子工业。| 黄,安德鲁。分类:LCC TK7836 .H83 2017(印刷版)| LCC TK7836(电子书)| DDC 621.381092--dc23 LC 记录可在 https://lccn.loc.gov/2016038846 获得
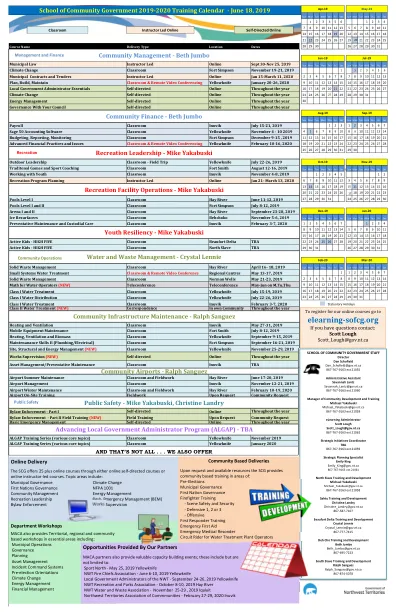
SCG_Training_Calendar_2019_2020 年 6 月 18 日.xlsx
MACA 合作伙伴还提供宝贵的能力建设活动;这些包括但不限于:Sport North —— 2019 年 5 月 25 日;黄刀市西北地区消防局长协会 —— 2019 年 6 月 6 日至 10 日;黄刀市西北地区地方政府管理员 —— 2019 年 9 月 24 日至 26 日;黄刀市西北地区休闲及公园协会 —— 2019 年 10 月 8 日至 10 日;海河西北地区水务及废物协会 —— 2019 年 11 月 25 日至 29 日;伊魁特西北地区社区协会 —— 2020 年 2 月 27 日至 29 日;因纽维克

使用 Oxford Nanopore 和 Illumina 平台对亚麻品种 Atlant 进行基因组测序
自古以来,人们就种植亚麻 ( Linum usitatissimum L. ) 以获取种子和纤维 ( Vaisey-Genser 和 Morris,2003 年 )。纤维亚麻比亚麻籽高,仅在茎的上部有分枝。亚麻籽的分枝从茎的中部开始,这些植物会产生许多大种子 ( Diederichsen 和 Richards,2003 年 )。亚麻籽富含 omega-3 脂肪酸和木脂素,其健康益处已在许多研究中得到证实 ( Caligiuri 等人,2014 年;Goyal 等人,2014 年;Kezimana 等人,2018 年;Parikh 等人,2019 年 )。因此,亚麻籽被用于食品和制药工业、动物饲料以及环保涂料和复合材料的生产(Singh 等人,2011;Corino 等人,2014;Goyal 等人,2014;Campos 等人,2019;Fombuena 等人,2019)。亚麻纤维是主要由纤维素组成的空心管;它们具有高强度和耐久性,可用于生产高质量的纺织品(Vaisey-Genser 和 Morris,2003)。亚麻纤维由于表面的芯吸和水分移动而具有很高的吸水能力,可用于制作炎热气候下的布料、帆、帐篷和地毯(Atton,1989)。然而,只有从亚麻茎的没有分支的部分才能获得长纤维;因此,尽管亚麻纤维质量很高,但它在很大程度上已被合成纤维所取代 ( Muir 和 Westcott,2003 年)。然而,对生态问题的认识引起了人们对使用对地球更具可持续性的材料的关注,人们对亚麻纤维的兴趣正在重新燃起。此外,在过去几年中,亚麻纤维已被积极用作复合材料的组成部分,在汽车、航空航天和包装应用中具有良好的潜力,在这些应用中,纤维长度并不十分重要 ( Zhu 等人,2013 年;Mokhothu 和 John,2015 年;Wu 等人,2016 年;Dhakal 和 Sain,2019 年;Fombuena 等人,2019 年;Goudenhooft 等人,2019 年;Zhang 等人,2020 年 a)。 2012 年,亚麻品种 CDC Bethune 的基因组在 Illumina 平台上进行了测序,采用双端和配对文库。结果组装结果为 302 Mb,其中 scaffild N50 约为 700 kb,contig N50 约为 20 kb,亚麻基因组覆盖率估计为 370 Mb,为 81%(Wang et al., 2012)。15 对 CDC 染色体的染色体水平组装

兽医学院名称:Sherein Salem
Engy M. Akl,Rasha S. Mohamed,Sherein S. Abdelgayed,Karem Fouda B,Mosaad A. Abdel-Wahhab D,“亚麻籽粘液的表征和抗氧化活性,评估其饮食中的饮食补充剂在改善钙的饮食补充剂中,可改善含量的100含量”。 https://doi.org/10.1016/j.bcdf.2024.100444

李叶荣、刘毅人、黄云。 2018.VicSim:通过情感和语言保真度增强受害者模拟。诉讼中
场景 用户担心楼上传来的巨响,他们认为那是他们大楼的二楼。用户提供的位置是“庭院公寓”,并分享了有关噪音的详细信息,包括噪音来自他们上方,并且发生在夜间和下午。用户还提到他们是加州大学洛杉矶分校的学生,他们没有向大学报告噪音。调度员派了一名警官到该地点,用户与警官 Jane 进行了交谈。警官确认噪音来自楼上,他们会进一步调查。序数:第二次时间:夜间时间:下午头衔:学生组织:加州大学洛杉矶分校人员:Jane

通过基因组编辑稻米核孔蛋白基因 OsCPR5.1(而非 OsCPR5.2)实现对水稻黄斑驳病毒的抗性
摘要 水稻黄斑驳病毒 (RYMV) 是非洲最严重的水稻疾病之一。RYMV 的管理具有挑战性。遗传抗性提供了最有效和最环保的控制。隐性抗性基因座 rymv2 (OsCPR5.1) 已在非洲水稻 (Oryza glaberrima) 中被鉴定,然而,渗入 Oryza sativa ssp。由于跨越障碍,粳稻和印度稻仍然具有挑战性。在这里,我们评估了两种水稻核孔蛋白旁系同源物 OsCPR5.1 (RYMV2) 和 OsCPR5.2 的 CRISPR/Cas9 基因组编辑是否可用于将 RYMV 抗性引入粳稻品种 Kitaake。两种旁系同源物均已被证实可弥补拟南芥 atcpr5 突变体的缺陷,表明存在部分冗余。尽管两种旁系同源物之间存在惊人的序列和结构相似性,但只有 o scpr5.1 功能丧失突变体完全具有抗性,而 oscpr5.2 功能丧失突变体仍然易感,这表明 OsCPR5.1 在 RYMV 易感性中起着特殊作用。值得注意的是,在 OsCPR5.1 的 N 端结构域(预计为非结构化)中存在短的框内删除或替换的编辑线对 RYMV 高度敏感。与单个拟南芥 AtCPR5 基因突变导致植物严重矮化不同,oscpr5.1 和 oscpr5.2 单敲除和双敲除突变体既没有表现出明显的生长缺陷,也没有表现出类似病变表型的症状,这可能反映了功能分化。OsCPR5.1 的特定编辑,同时保持 OsCPR5.2 活性,为在优良稻种系中产生 RYMV 抗性以及与其他 RYMV 抗性基因或其他性状有效叠加提供了一种有前途的策略。