XiaoMi-AI文件搜索系统
World File Search SystemA05
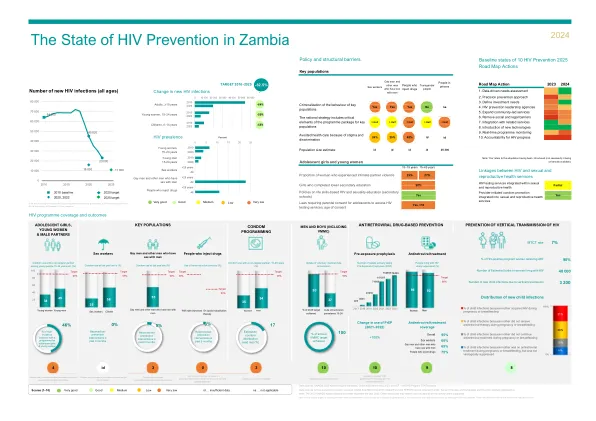
2024 年全球艾滋病预防记分卡最终版 a05 (002).xlsx
非常好 良好 中 低 非常低 id … 数据不足 na … 不适用 重点人口计划覆盖范围的数据来源:2023 年全球艾滋病监测、全球基金和 PEPFAR 于 2023 年获得的报告。部分数据是三角测量的,因此不具有全国代表性。
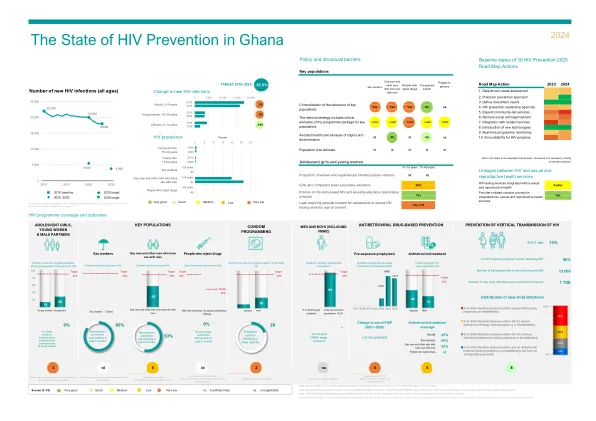
2024 年全球艾滋病预防记分卡最终版 a05 (002).xlsx
非常好 良好 中 低 非常低 id … 数据不足 na … 不适用 重点人口计划覆盖范围的数据来源:2023 年全球艾滋病监测、全球基金和 PEPFAR 于 2023 年获得的报告。部分数据是三角测量的,因此不具有全国代表性。
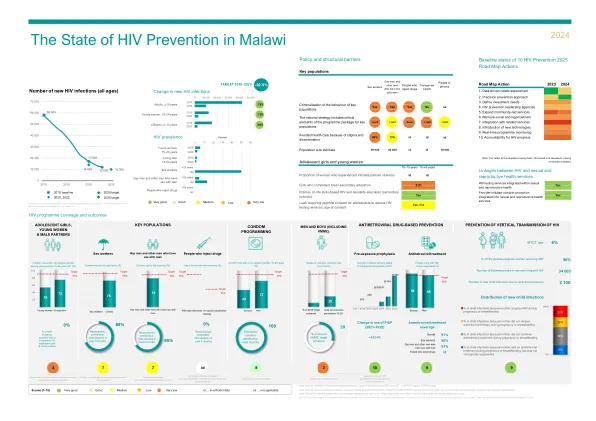
2024 年全球艾滋病预防记分卡最终版 a05 (002).xlsx
非常好 良好 中 低 非常低 id … 数据不足 na … 不适用 重点人口计划覆盖范围的数据来源:2023 年全球艾滋病监测、全球基金和 PEPFAR 于 2023 年获得的报告。部分数据是三角测量的,因此不具有全国代表性。

无标题 - 国际核信息系统 (INIS)
j(^ii>'adV'.Si>ring'ne^d.. ' \'A 2.2X&1..NTIS pi-iere'codes;Npri ; ti tt'd 'copy — A05, ni:ii:ro'fic 1 iie*-c*o.ijy ^~ .v|j.l。这个 .repqi-t 被准备为 n-n 。'。vt Q'f^wo^'k" 由 tJl.e- Vnited'.St Ues IGovei'nriien't 的 a'iV 机构赞助^t'ed。L'hited |5t 都不是吗?ate.s Gbv.^rrimt'iit 'tioi'。任何机构 • rf t i'-ni5i\'any “Of,-their雇佣^©St-nor-.any : t)f;theit'C ?pnt '演员。s'tilH'phtractovs," 或他们的 'einpiQye.0^, niak'ejs: 任何 v .warranty.,, exp'^es^ !Bt'd,;'oi:;iissUmes 任何 li'gal-责任- br.respeiosibili't-y ft >•:准确性..完整性,或riijj的使用| ijifornia'硫醇装置',' i|i,.. ipfHrigt'p'i'tvajel.v ow'ned .rig :&|si。 Refevi?n,CL>••此处为 iiny 'spetifi.e'' ^f^ia:i;pVod;uetJ.i)rocessyQ.r;s(Si'^ 制造商,或' : pthe<' \vis.e,-dQ®.lipt%necei^ari:iy 。 epristitute oj;注射;'ipfHrigt'p'i'tvajel.v ow'ned .rig :&|si。Refevi?n,CL>••此处为 iiny 'spetifi.e'' ^f^ia:i;pVod;uetJ.i)rocessyQ.r;s(Si'^ 制造商,或' : pthe<' \vis.e,-dQ®.lipt%necei^ari:iyepristitute oj;注射;'

流体动力学分部 - APS DFD 2022
上午 8:00 – 上午 9:57 并行会议 A01 焦点会议:流体 接下来:软体撞击流体 I Sagamore 宴会厅 1–7 A02 空气动力学:常规 130 A03 主动物质 I:主动湍流 131 A04 动脉瘤 132 A05 动物飞行:飞行昆虫 I 133 A06 高雷诺数游泳 I 134 A07 生理、发声和言语 135 A08 气泡:常规 136 A09 CFD:浸入边界法 I 137 A10 粒子-湍流相互作用 I 138 A11 声学:常规 139 A12 颗粒流 I 140 A13 生物流体动力学:生理 I 141 A14 自由表面流:常规142 A15 实验技术:生物和多相测量 143 A16 流动控制:概述 144 A17 流动不稳定性:多相流和瑞利-泰勒 145 A18 喷射流 I 205 A19 非牛顿流:理论与建模 206 A20 非线性动力学:库普曼和相关方法 207 A21 湍流:湍流建模的机器学习方法 I 208 A22 多孔介质流:对流和传热 231 A23 自由表面流:自然流 232 A24 反应流:LES 和 DNS 233 A25 表面张力效应:界面现象 I 234 A26 波:非线性动力学与湍流 235 A27 涡旋动力学:概述 I 236 A28 CFD:不确定性量化和机器学习 237 A29 液滴:电场效应 238 A30 液滴:超疏水表面和多液滴相互作用 239 A31 流动不稳定性:复杂流体 240 A32 地球物理流体动力学:大气 241 A33 微/纳米流动:通道 242 A34 相变 I 243 A35 一般流体动力学:越过障碍物的流动 244
无标题 - 国际核信息系统 (INIS)
jiefitsf 能量.. BropkhiUjen 是;一个肯定的 aciic n/平等机会雇主'...报告'.Vb, BNL pS!3(>0.bpE^QSTl-45()0-- fSt^.^iiitt'ibution caJeg.oiii<|s'L'C~tO() a.nd C.C-.50.0.^ gt fieivdl iiviscc'tlanebusand;'过程ro^jqtts |iiuclearandTibri.iiut'leaiM。Pi'inte.tf ', "t'nited'St'ateisirfsVn'Vrieai^vaiUibiefi'bm Strvict'., VjS..J?t r pft.i'tni'.cnt|of 1 t'Q'n >nit'rpt''528S.端口 j(^ii>'adV'.Si>ring'ne^d.. ' \'A 2.2X&1..NTIS pi-iere'codes;Npri; ti tt'd '复制 — A05,ni:ii:ro'fic 1 iie*-c*o.ijy ^~ .v|j.l.此 .repqi-t 已准备好为 n-n .'。该项目由圣彼得堡国立大学的一个机构赞助。L'hited 也不 |5t ?ate.s Gbv.^rrimt'iit 'tioi'。任何机构 • rf t i'-ni5i\'any “Of,-their employ^©St-nor-.any : t)f;theit'C ?pnt'演员。s'tilH'phtractovs”,或其“einpiQye.0^,niak'ejs:任何 v .warranty.,, exp'^es^ !Bt'd,;'oi:;iissUmes 任何 li'gal-责任范围包括:设备的准确性、完整性或使用性,包括但不限于: ; bi:r,epfes ipfHrigt'p'i'tvajel.v ow'ned .rig : &|si. 参考,CL>••此处指任何“特定”^f^ia:i;pVod;uetJ.i)rocessyQ.r;s(Si'^制造商,或' : pthe<' \vis.e,-dQ®.lipt%necei^ari:iy . epristitute oj;注入;' .iQj^e»^:t/'-i:tc6ift'mi! 'ricfeli«rt5'OT!-fa.yoi;in 1 ^ b.v;the. tijiift d'Statbs.Gp.vernm.ftrit oi' &ny' i 'agen-e^t'vw^ffl^iftioV'br%u'bfqn'tKai! 't^t''fh'ifti'C(itfc' - --ipfHrigt'p'i'tvajel.v ow'ned .rig : &|si.参考,CL>••此处指任何“特定”^f^ia:i;pVod;uetJ.i)rocessyQ.r;s(Si'^制造商,或' : pthe<' \vis.e,-dQ®.lipt%necei^ari:iyepristitute oj;注入;' .iQj^e»^:t/'-i:tc6ift'mi!'ricfeli«rt5'OT!-fa.yoi;in 1 ^ b.v;the.tijiift d'Statbs.Gp.vernm.ftrit oi' &ny' i 'agen-e^t'vw^ffl^iftioV'br%u'bfqn'tKai!'t^t''fh'ifti'C(itfc' - --

来自种间甘蓝rapa衍生物的新型定量性状基因座改善了甘蓝纳普斯napus
POD破碎是农业相关性的一种特征,可确保植物在其本地环境中取代种子,并在几种宽阔的农作物中受到了驯化和选择的驯化和选择。然而,豆荚破碎会导致菜籽(甘蓝纳普斯L.)作物的显着屈服降低。衍生自B. rapa/b的种间繁殖线BC95042。Napus Cross表现出改善的POD破碎阻力(比易碎的B. Napus品种高达12倍)。为了揭示新品种中的遗传基础并改善了POD破碎的耐药性,我们分析了F 2和F 2:3衍生的种群,来自BC95042和Advanced Breeding系列的交叉,BC95041,并用15,498 Dartseq标记的基因分型。通过基因组扫描,间隔和包容性的复合间隔映射分析,我们确定了与POD破裂能量相关的七个定量性状基因座(QTL),用于POD破碎的抗性或POD强度的度量,并且它们位于A02,A02,A03,A03,A05,A09,A09,A09和C01 Chromosomes上。两种亲本线都为豆荚碎片抗性贡献了等位基因。我们确定了添加剂X添加剂,添加性优势和优势X优势X在A01/C01,A01/C01,A03/A07,A07/C03,A03,A03/C03和C01/C02染色体之间的相互作用之间的五对X添加剂,添加剂优势和优势X优势相互作用。QTL对A03/ A07和A01/ C01的影响处于排斥阶段。比较映射确定了几种候选基因(AG,ABI3,BP1,CEL6,FIL,FIL,FUL,GA2OX2,IND,LATE,LEUNIG,MAGL15,RPL,QRT2,RGA,RGA,SPT,SPT和TCP10),基于QTL和QTL的QTL和上毒QTL相互作用,以实现pod shatter pod shatter shatter shatter shatter shatter shatter shatter shatters。BNAA09G05500D受到在A02,A03和A09上检测到的三个QTL靠近(富有成果的)同源物BNAA03G39820D和BNAAA09G05500D。着眼于FUL,我们研究了推定的图案,序列变体和其同源物的进化速率,373个重新设备的B. napus napus感兴趣。
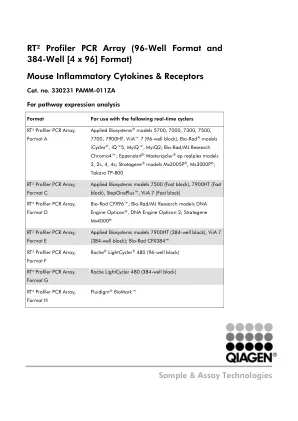
RT² Profiler PCR 阵列(96 孔格式和 384 ... - GeneGlobe
A01 Mm.235137 NM_007926 Aimp1 氨酰 tRNA 合成酶复合物相互作用多功能蛋白 1 A02 Mm.103205 NM_007553 Bmp2 骨形态发生蛋白 2 A03 Mm.1283 NM_011329 Ccl1 趋化因子(CC 基序)配体 1 A04 Mm.4686 NM_011330 Ccl11 趋化因子(CC 基序)配体 11 A05 Mm.867 NM_011331 Ccl12 趋化因子(CC 基序)配体 12 A06 Mm.41988 NM_011332 Ccl17 趋化因子(CC 基序)配体 17 A07 Mm.424740 NM_011888 Ccl19 趋化因子(CC 基序)配体 19 A08 Mm.290320 NM_011333 Ccl2 趋化因子(CC 基序)配体 2 A09 Mm.116739 NM_016960 Ccl20 趋化因子(CC 基序)配体 20 A10 Mm.12895 NM_009137 Ccl22 趋化因子(CC 基序)配体 22 A11 Mm.31505 NM_019577 Ccl24 趋化因子(CC 基序)配体 24 A12 Mm.1282 NM_011337 Ccl3 趋化因子(CC 基序)配体 3 B01 Mm.244263 NM_013652 Ccl4 趋化因子(CC 基序)配体 4 B02 Mm.284248 NM_013653 Ccl5 趋化因子(CC 基序)配体 5 B03 Mm.137 NM_009139 Ccl6 趋化因子(CC 基序)配体 6 B04 Mm.341574 NM_013654 Ccl7 趋化因子(CC 基序)配体 7 B05 Mm.42029 NM_021443 Ccl8 趋化因子(CC 基序)配体 8 B06 Mm.416125 NM_011338 Ccl9 趋化因子(CC 基序)配体 9 B07 Mm.274927 NM_009912 Ccr1 趋化因子(CC 基序) 受体 1 B08 Mm.8021 NM_007721 Ccr10 趋化因子 (CC 基序) 受体 10 B09 Mm.6272 NM_009915 Ccr2 趋化因子 (CC 基序) 受体 2 B10 Mm.57050 NM_009914 Ccr3 趋化因子 (CC 基序) 受体 3 B11 Mm.1337 NM_009916 Ccr4 趋化因子 (CC 基序) 受体 4 B12 Mm.14302 NM_009917 Ccr5 趋化因子 (CC 基序) 受体 5 C01 Mm.8007 NM_009835 Ccr6 趋化因子 (CC 基序) 受体 6 C02 Mm.442098 NM_007720 Ccr8 趋化因子(CC 基序)受体 8 C03 Mm.4861 NM_011616 Cd40lg CD40 配体 C04 Mm.795 NM_007778 Csf1 集落刺激因子 1(巨噬细胞) C05 Mm.4922 NM_009969 Csf2 集落刺激因子 2(粒细胞-巨噬细胞) C06 Mm.1238 NM_009971 Csf3 集落刺激因子 3(粒细胞) C07 Mm.103711 NM_009142 Cx3cl1 趋化因子(C-X3-C 基序)配体 1 C08 Mm.21013 NM_008176 Cxcl1 趋化因子(CXC 基序)配体 1 C09 Mm.877 NM_021274 Cxcl10 趋化因子(CXC 基序)配体 10 C10 Mm.131723 NM_019494 Cxcl11 趋化因子(CXC 基序)配体 11 C11 Mm.303231 NM_021704 Cxcl12 趋化因子(CXC 基序)配体 12 C12 Mm.10116 NM_018866 Cxcl13 趋化因子(CXC 基序)配体 13 D01 Mm.64326 NM_011339 Cxcl15 趋化因子(CXC 基序)配体 15 D02 Mm.4660 NM_009141 Cxcl5 趋化因子(CXC 基序)配体 5 D03 Mm.766 NM_008599 Cxcl9 趋化因子(CXC 基序)配体 9 D04 Mm.234466 NM_009909 Cxcr2 趋化因子(CXC 基序)受体 2 D05 Mm.12876 NM_009910 Cxcr3 趋化因子(CXC 基序)受体 3 D06 Mm.6246 NM_007551 Cxcr5 趋化因子(CXC 基序)受体 5 D07 Mm.3355 NM_010177 Fasl Fas 配体(TNF 超家族,成员 6) D08 Mm.240327 NM_008337 Ifng 干扰素伽马 D09 Mm.379327 NM_008348 Il10ra 白细胞介素10 受体,α