XiaoMi-AI文件搜索系统
World File Search SystemMYCN

OncoTrack 液体
ABL1、ABL2、AKT1、ALK、APC、AR、ARAF、ARID1A、ARID1B、ATM、ATR、ATRX、BAP1、BARD1、BRAF、BRCA1、BRCA2、BRIP1、C11orf65、CCND1、CDH1、CDK12、CDK4、CDKN2A、CDX2、CHEK1、CHEK2、CSF1R、CTNNB1、DDR2、EGFR、ERBB2、ERBB3、ERBB4、ERCC2、ESR1、EZH2、FANCL、FBXW7、FGFR1、FGFR2、FGFR3、FGFR4、FLT3、FOXA1、FOXL2、GATA3、GNA11、GNAQ、GNAS、HNF1A、HRAS、IDH1、IDH2、INPP4B、 JAK1、JAK2、JAK3、KDM5C、KDM6A、KEAP1、KIT、KRAS、MAP2K1、MAP2K2、MAPK1、MET(外显子 14 跳跃)突变)、MLH1、MPL、MSH2、MSH6、MTOR、MUTYH、MYC、MYCN、MYD88、NF1、NF2、NOTCH1、NPM1、NRAS、NTRK1、NTRK3、PALB2、PBRM1、PDGFRA、PIK3CA、PMS2、POLD1、POLE、PPP2R2A、PTCH1、PTEN、PTPN11、RAD51B、RAD51C、RAD51D、RAD54L、RAF1、RB1、RET、RHEB、RHOA、RIT1、ROS1、SETD2、SF3B1、 SMAD4、SMARCB1、SMO、SPOP、SRC、STK11、TERT、TP53、TSC1、TSC2、VHL

肿瘤休眠与神经母细胞瘤的预后预测和肿瘤免疫密切相关
神经母细胞瘤(NB)是童年时期最常见,最致命的实体瘤,其特征是从自发性回归到无情进展的异质行为(1)。尽管早期诊断和新的治疗方法,但NB复发仍然是该领域最大的临床挑战之一,并对生存产生了不利影响(2)。在大量的NB儿童中,遥远的转移发生了多年甚至数十年的潜伏期。在50%以上的病例中发生复发,5年生存率小于40%(3)。许多生物标志物已向医师提供有关NB的治疗和预后的指导,包括国际神经母细胞瘤分期系统(INSS)阶段,MYCN(N-MYC,骨髓细胞膜症癌),诊断年龄,诊断年龄,组织学类别,分化等级,差异,Karyorrhexis Inderex(Mkaryorrhexis Inder)(Mkaryorrhexis)(Mkiy)(Mkiy)(Mki)和pna。但是,这些生物标志物都无法准确预测复发(4,5)。尽管在强化多模式治疗方面取得了进展,但大多数高风险NB诊断时会出现广泛转移性疾病,并且在治疗后最初或最终对常规疗法反应,长期生存率小于50%(3)。免疫疗法可以大大提高癌症患者的存活率,而
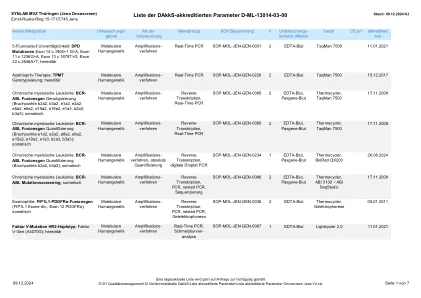
认可参数列表 Oncoscreen Jena v5.xlsx
Oncomine Comprehensive Assay v3 DNA 组:AKT1、AKT2、AKT3、ALK、AR、ARAF、ARID1A、ATM、ATR、ATRX、AXL、BAP1、BRAF、BRCA1、BRCA2、BTK、CBL、CCND1、CCND2、CCND3、CCNE1、CDK12、CDK2、CDK4、CDK6、CDKN1B、CDKN2A、CDKN2B、CHEK1、CHEK2、CREBBP、CSF1R、CTNNB1、DDR2、EGFR、ERBB2、ERBB3、ERBB4、ERCC2、ESR1、EZH2、FANCA、FANCD2、FANCI、FBXW7、FGF19、FGF3、FGFR1、FGFR2、FGFR3、FGFR4、FLT3、 FOXL2、GATA2、GNA11、GNAQ、GNAS、H3-3A、HIST1H1E、HNF1A、HRAS、IDH1、IDH2、IGF1R、JAK1、JAK2、JAK3、KDR、KIT、KNSTRN、KRAS、MAGOH、MAP2K1、MAP2K2、MAP2K4、MAPK1、MAX、MDM2、 MDM4、MED12、MET、MLH1、MRE11A、MSH2、MSH6、MTOR、MYC、MYCL、MYCN、MYD88、NBN、NF1、NF2、NFE2L2、NOTCH1、NOTCH2、NOTCH3、NRAS、NTRK1、NTRK2、NTRK3、PALB2、PDGFRA、PDGFRB、PIK3CA、 PIK3CB, PIK3R1、PMS2、POLE、PPARG、PPP2R1A、PTCH1、PTEN、PTPN11、RAC1、RAD50、RAD51、RAD51B、RAD51C、RAD51D、RAF1、RB1、RET、RHEB、RHOA、RICTOR、RNF43、ROS1、SETD2、SF3B1、SLX4、SMAD4、SMARCA4、SMARCB1、SMO、SPOP、SRC、STAT3、STK11、TERT、TOP1、TP53、TSC1、TSC2、U2AF1、XPO1

研究论文比较神经母细胞瘤细胞中 RIST 分子靶向方案中的 mTOR 抑制剂雷帕霉素和 Torin-2
复发或难治性高危神经母细胞瘤患者的预后仍然不容乐观,迫切需要新的治疗方案。RIST 治疗方案采用多模式节拍疗法设计,结合分子靶向药物(雷帕霉素和达沙替尼)和化疗主干(伊立替康和替莫唑胺),目前正在 II 期临床试验(NCT01467986)中验证。随着新型、更有效的 ATP 竞争性 mTOR 抑制剂的出现,我们期望改进 RIST 联合疗法。通过比较 Torin-1、Torin-2、AZD3147 和 PP242 的 IC 50 值,我们确定只有 Torin-2 抑制了在纳摩尔浓度下测试的所有三种 MycN 扩增神经母细胞瘤细胞系的细胞活力。两种 mTOR 抑制剂的单独治疗诱导了显著的 G 1 细胞周期停滞,与达沙替尼联合治疗降低了细胞周期调节剂细胞周期蛋白 D1 的表达或增加了细胞周期抑制剂 p21 的表达。两种 mTOR 抑制剂的组合指数表明与达沙替尼具有协同作用。有趣的是,与雷帕霉素相比,与 Torin-2 联合治疗可产生更广泛的 mTOR 通路抑制,这表现为 AKT(Thr308、Ser473)、4E-BP(Ser65)和 S6K(Thr389)磷酸化降低。此外,在改良的多模态 RIST 方案中用 Torin-2 替代雷帕霉素可降低细胞活力并诱导细胞凋亡,尽管所用的 Torin-2 药物浓度明显较低。纳摩尔浓度的功效可显著降低与雷帕霉素相关的不必要的免疫抑制。然而,目前我们不能排除 Torin-2 因其在更复杂系统中的效力而增加毒性的可能性。尽管如此,我们的结果表明,在 RIST 方案中加入 Torin-2 作为雷帕霉素的替代品可能是一种有效的选择,可以在复发或治疗难治性高危神经母细胞瘤的前瞻性临床试验中进行评估。

与IRF2BPL基因变异的新型人类神经发育和神经退行性疾病 - 机械和治疗途径
确定新型的治疗方法,该方法利用了特定的肿瘤脆弱性。与成年癌症相比,通常表现出一生中积累的大量突变,小儿肿瘤通常在组织范围内的发育窗口中出现 - 特定方式 - 通常只有很少的突变驱动因素和低突变负担(4)。小儿实体瘤中的一个共同特征是融合癌蛋白的存在,由于染色体畸变而出现了(5)。此外,在某些儿科实体瘤中频繁进行肉体内和外肿瘤性癌基因的扩增,例如在神经母细胞瘤中,在神经母细胞瘤中,经常在ECDNA上发生myCN扩增,这是对不良预后的预测因子(6-10)。基因扩增和融合癌蛋白都难以直接治疗,尤其是在影响转录因子时,这阻碍了这些肿瘤实体中选择性疗法的发展。基因组不稳定性是癌细胞的标志(11),最近已证明它在治疗上可起作用(12)。癌细胞中的极端增殖率部分由融合型癌蛋白和癌基因扩增引起,可能会导致所谓的复制应力的DNA延迟或误差(13-15)。响应受损的DNA,细胞具有复杂的机制来识别和修复病变,同时确保细胞周期停止,称为DNA损伤响应(DDR)。DDR主要由三种激酶调节:共济失调突变(ATM),共济失调telangiectasia-和rad3相关(ATR)以及DNA依赖性蛋白激酶催化亚基(DNA-PKC; ref。16)。即使它们具有相似的蛋白质序列,并且它们的靶标重叠,但它们对它们对不同刺激的反应也被广泛接受(17)。尽管ATM和DNA-PKC在双链断裂后大部分被激活,但ATR主要响应复制应力与与DNA相关的DNA损伤,这通常涉及单链DNA中间体(18、19)。由于ATR响应于复制应力而被激活,因此有人提出,癌症比非转化的细胞更强烈地依赖于ATR来耐受高水平的复制应力(20,21)。这些发现激发了测试ATR抑制剂作为癌症治疗选择的兴趣,尤其是在具有较高复制应激的肿瘤中。一些预测的生物标志物

pdf 14p3439.pdf
原理:代谢的协同重编程主导神经母细胞瘤 (NB) 的进展。基于阐明代谢重编程的分子机制,开发一种具有分层指导的 NB 治疗选择的个性化风险预测方法具有重要的临床意义。方法:利用基于机器学习的多步骤程序,在单细胞和代谢物通量维度上阐明代谢重编程驱动的 NB 恶性进展的协同机制。随后,开发了一种有前景的代谢重编程相关预后特征 (MPS) 和基于 MPS 分层的个性化治疗方法,并使用临床前模型进一步独立验证。结果:MPS 鉴定的 MPS-I NB 表现出比 MPS-II 对应物明显更高的代谢重编程活性。 MPS 在预测预后方面比目前的临床特征 [AUC:0.915 vs. 0.657(MYCN)、0.713(INSS 分期)和 0.808(INRG 分层)] 表现出更高的准确性。AZD7762 和依托泊苷分别被确定为针对 MPS-I 和 II NB 的有效治疗药物。后续生物学测试表明,AZD7762 显著抑制 MPS-I NB 细胞的生长、迁移和侵袭,且效果优于 MPS-II 细胞。相反,依托泊苷对 MPS-II NB 细胞的治疗效果更好。更令人鼓舞的是,AZD7762 和依托泊苷分别显着抑制了 MPS-I 和 MPS-II 样本中的体内皮下肿瘤形成、增殖和肺转移;从而延长了荷瘤小鼠的生存期。从机制上看,AZD7762 和依托泊苷分别通过线粒体依赖性途径诱导 MPS-I 和 MPS-II 细胞凋亡;而 MPS-I NB 通过依赖谷氨酸代谢和乙酰辅酶 A 抵抗依托泊苷诱导的细胞凋亡。MPS-I NB 进展受到多种代谢重编程驱动因素的推动,包括多药耐药性、免疫抑制和促肿瘤炎症微环境。从免疫学上看,MPS-I NB 通过 MIF 和 THBS 信号通路抑制免疫细胞。代谢方面,重编程的谷氨酸代谢、三羧酸循环、尿素循环等显著支持了 MPS-I NB 细胞的恶性增殖。此外,MPS-I NB 细胞表现出独特的促肿瘤发育谱系和自我通讯模式,这表现为随着发育和自我通讯而激活的致癌信号通路增强。结论:本研究深入了解了代谢重编程介导的 NB 恶性进展的分子机制。它还为开发以新的精确风险预测方法为指导的靶向药物提供了启示,这可能有助于显著改善 NB 的治疗策略。