XiaoMi-AI文件搜索系统
World File Search SystemNR4A

成功产生了具有强大抗肿瘤作用的CAR-T细胞
[图2]在与癌细胞相同的培养基中培养CAR-T细胞时,用于耗尽CAR-T细胞的实验模型,必须将CAR-T细胞移至每3-4天含有新癌细胞的培养基,因为CAR-T细胞杀死了癌细胞。 CAR-T细胞是由五个健康供体产生的,并比较了破坏三个NR4A基因的野生型CAR-T细胞(红色)和CAR-T细胞(蓝色)的数量。尽管在培养后,来自任何供体的野生型CAR-T细胞都耗尽了大约14天,但停止了细胞增殖,但缺乏NR4A的CAR-T细胞继续增殖。

天然产物作为抗癌剂并通过基于机制的精准方法增强其功效
传统药物及其活性成分以及一些天然产物和衍生类似物已被用于治疗多种疾病,包括癌症。在这些化合物中,细胞毒性剂如博来霉素、紫杉醇和长春新碱可阻断癌细胞生长所需的基本途径和基因,这些药物具有多种临床应用。膳食酚类化合物(包括黄酮类化合物和相关化合物)具有多种健康益处,然而,大多数在临床前研究中显示出良好抗癌活性的个体膳食化合物和其他天然产物的临床效果极小,对癌症尤其如此。由于药代动力学考虑和摄取有限(例如姜黄素),许多化合物在临床试验中表现不佳,这些问题是可以解决的。黄酮类化合物和许多其他天然产物衍生的抗癌化合物的临床效果也可以通过更有针对性的方法得到增强。这将包括识别特定癌症中的显著反应/基因或靶点,然后确定最佳化合物。在这篇评论中,我讨论了有限数量的靶标,包括非致癌基因成瘾基因,例如 Sp 转录因子、活性氧 (ROS) 或孤儿核受体 4A (NR4A) 亚家族。因此,对这些反应最有效的化合物只能用于治疗 ROS 诱导或高表达靶标(例如 Sp1 或 NR4A 亚家族成员)的患者。基于机制的精准医疗方法应能提高饮食和相关天然产品作为抗癌剂的临床疗效,并减少某些联合疗法的毒副作用。

feng 2020-0052-ms_cn
摘要:NUR77属于核受体超家族的NR4A亚组。与其他核受体不同,尚未确定NUR77的天然配体。但是,一些小分子可以与该受体相互作用,并诱导构象变化以介导其活性。使用各种生理和病理刺激可以快速增加NUR77的表达和激活。体内和体外研究表明,通过参与细胞分化,凋亡,代谢,线粒体稳态和其他过程,其在多个系统的组织和细胞中的调节作用。尽管目前在中枢神经系统(CNS)的病理生理学中对NUR77的研究受到限制,但目前的数据支持了NUR77参与许多神经系统疾病,例如中风,多发性硬化症,帕金森氏病。这表明NUR77的激活在治疗这些疾病方面具有很大的潜力。本综述总结了中枢神经系统疾病中NUR77的调节机制,并为其作为靶向疗法的潜力提供了可用的证据,尤其是与脑血管和炎症相关的中枢神经系统疾病。
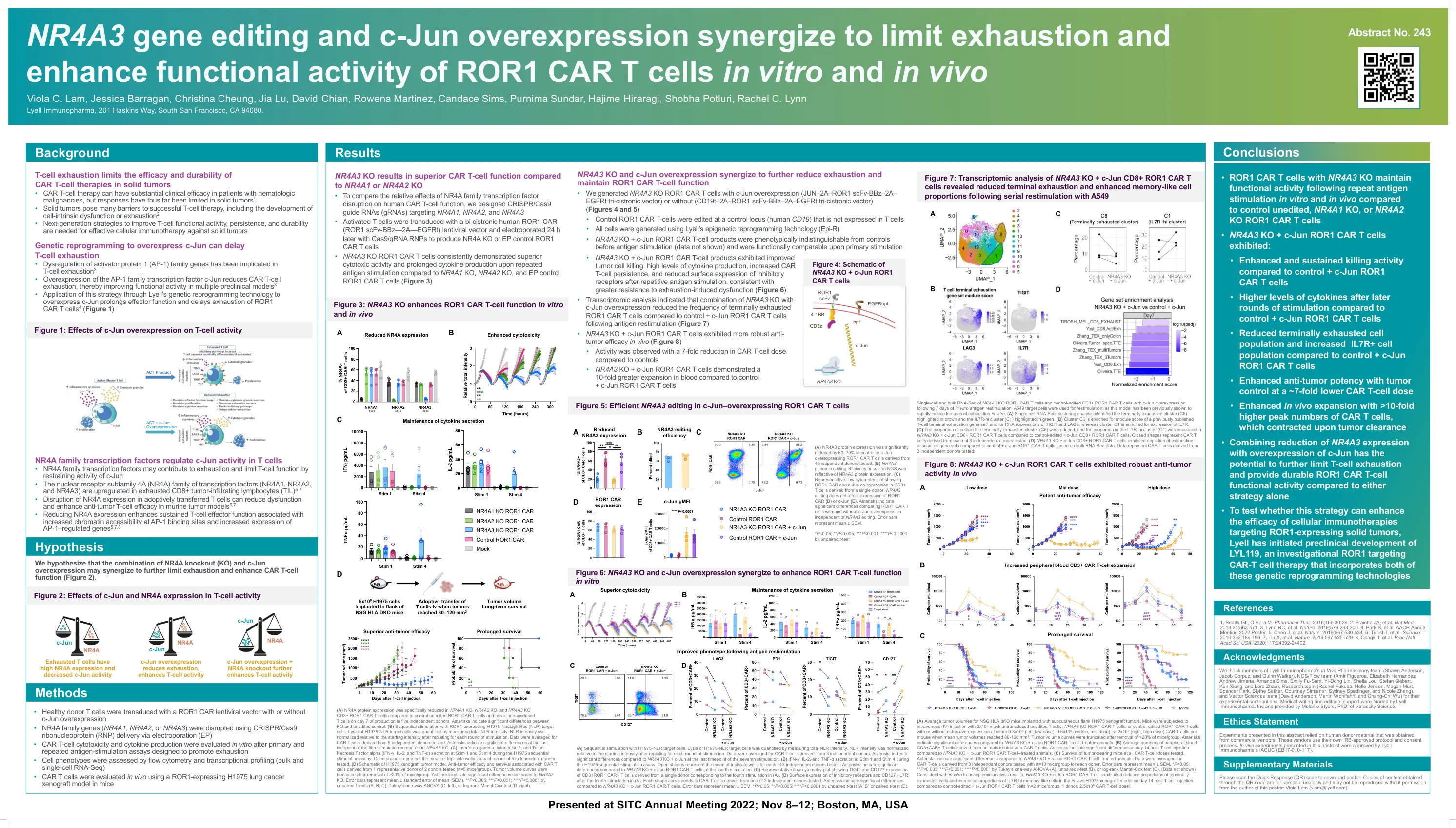
NR4A3 基因编辑和 c-Jun 过表达协同作用,限制 ROR1 CAR-T 细胞衰竭并增强其体内外功能活性
(A) 在 5 位独立供体中,与对照未编辑 ROR1 CAR T 细胞和模拟未转导 T 细胞相比,在生产第 7 天,NR4A1 KO、NR4A2 KO 和 NR4A3 KO CD3+ ROR1 CAR T 细胞中的 NR4A 蛋白表达明显降低。星号表示 KO 和未编辑对照之间存在显着差异。 (B) 用表达 ROR1 的 H1975-NucLightRed (NLR) 靶细胞进行顺序刺激。通过测量总 NLR 强度来量化 H1975-NLR 靶细胞的裂解。在每轮刺激后重新接种后,NLR 强度相对于起始强度进行标准化。对来自 5 位独立供体的 CAR T 细胞取平均值。星号表示与 NR4A3 KO 相比,第五次刺激的最后一个时间点存在显着差异。 (C) 在 H1975 顺序刺激试验期间,刺激 1 和刺激 4 时干扰素 γ、白细胞介素 2 和肿瘤坏死因子 α (IFN-γ、IL-2 和 TNF-α) 的分泌。空心形状表示对 5 个独立供体进行测试的每个供体的三重孔的平均值。 (D) H1975 异种移植肿瘤模型示意图。来自 2 个测试供体中的 1 个代表性供体的 CAR T 细胞的抗肿瘤功效和存活率(n=5 只小鼠/组)。在去除每组 >20% 的小鼠后,肿瘤体积曲线被截断。星号表示与 NR4A3 KO 相比有显着差异。误差线表示平均值±平均值的标准误差 (SEM)。** P <0.005;*** P <0.001; **** 非配对 t 检验 (A、B、C)、Tukey 单向方差分析 (D,左) 或对数秩 Manel-Cox 检验 (D,右) 的 P < 0.0001。