XiaoMi-AI文件搜索系统
World File Search SystemOXPHOS

烟酰胺单核苷酸(NMN)在2型糖尿病中通过脂肪组织中的意外作用,而不是通过线粒体生物发生
摘要:烟酰胺单核苷酸(NMN)已成为针对年龄相关疾病(包括2型糖尿病)的有希望的治疗干预措施。在这项研究中,我们证实了NMN治疗对葡萄糖摄取的先前观察到的影响,并研究了其在各种组织和细胞系中的基本机制。通过迄今为止最全面的蛋白质组学分析,我们发现了一系列新型器官特异性作用,负责通过IPGTT:脂肪组织生长来测量的葡萄糖摄取(通过增加蛋白质合成,降解和MTORPORPRIFTIAL SIGNAMENTION提高)。值得注意的是,我们观察到了热UCP1的上调,促进了肌间脂肪组织中的葡萄糖转化为热量,同时显示出对肌肉和大脑线粒体生物发生的令人惊讶的抑制作用。此外,肝脏和肌肉细胞表现出独特的反应,其特征是剪接体下调和同时上调伴侣,蛋白酶体和核糖体,从而导致轻度受损和能量感知的蛋白质合成机制。此外,我们的发现揭示了大脑中明显的代谢重新布线。这涉及增加酮体的产生,线粒体Oxphos和TCA循环成分的下调以及诱导众所周知的禁食相关作用。共同阐明了NMN作用的多方面性质,突出了其特异性效应及其在改善葡萄糖摄取中的作用。这些发现加深了我们对NMN治疗潜力的理解,并为管理代谢疾病的新策略铺平了道路。

癌症药物耐药性
摘要 恶性造血细胞获得代谢可塑性,重组合成代谢机制以提高合成代谢输出并防止氧化损伤,并绕过细胞周期检查点,最终胜过正常造血细胞。目前急性髓系白血病 (AML) 的治疗策略基于预后分层,其中包括突变谱作为最接近疾病生物学的替代指标。靶向治疗的临床疗效,例如针对突变型 FMS 样酪氨酸激酶 3 (FLT3) 和异柠檬酸脱氢酶 1 或 2 的药物,大多限于相关突变的存在。最近的研究不仅证明 AML 中的特定突变会造成代谢脆弱性,而且还强调了针对代谢脆弱性与这些突变抑制剂联合使用的功效。因此,阐明这些脆弱性的遗传分层、代谢依赖性和对特定抑制剂的反应之间的功能关系对于确定更有效的治疗方案、了解耐药机制和确定早期反应标记至关重要,最终提高治愈的可能性。此外,肿瘤微环境中发生的代谢变化也被报道为治疗靶点。白血病干细胞 (LSC) 的代谢特征不同,复发/难治性 LSC 会切换到替代代谢途径,促进氧化磷酸化 (OXPHOS),使其具有治疗耐药性。在这篇综述中,我们讨论了癌症代谢途径在 AML 代谢可塑性中的作用,并赋予对标准疗法的耐药性;我们还重点介绍了该领域在将这些重要发现转化为临床方面的最新有希望的进展,并讨论了支持代谢可塑性和与 AML 细胞相互作用的肿瘤微环境。

Nelarabine在T细胞急性淋巴细胞白血病
lon蛋白酶1(LONP1)是位于线粒体基质中的ATP依赖性蛋白酶,在调节线粒体蛋白抑制性,代谢和细胞应激反应等方面起着至关重要的作用。在各种肿瘤的进展中发现了异常的LONP1表达。然而,LONP1在前列腺癌(PCA)中的作用和分子机制仍然知之甚少。在这里我们表明,LONP1的过表达与PCA患者的不良临床病理特征和预后不良密切相关。机械上,发现发现LONP1与从氧化磷酸化(OXPHOS)转变为有氧糖酵解的代谢转换有关,从而促进肿瘤的增殖,侵袭和转移,并在体外和体内进行转移。同时,我们证明LONP1作为蛋白酶直接靶向线粒体丙酮酸载体1(MPC1),这是一种在糖酵解过程中的关键代谢蛋白,并增强其降解,从而又抑制了三羧酸(TCA)周期,并最终促进PCA的进展。Furthermore, using PCa in cancer-prone mice homozygous for a prostate-targeted conditional Pten knockout and Lonp1 knockin, we integrate transcriptomic and proteomic analyses of prostate tumors, upon which reveals that Lonp1 overexpression results in a signi fi cant downregulation of NADH: ubiquinone oxidoreductase activity, consequently impeding the electron transfer process and线粒体ATP合成,与PCA转移有关。总的来说,我们的结果表明,PCA中LONP1引起的代谢重编程与疾病进展紧密相结合,这表明针对线粒体中LONP1介导的级联反应可能会为PCA疾病提供治疗潜力。

隐性MECR致病变异引起LHON
抽象背景Leber的遗传性视神经神经病(LHON)是一种线粒体疾病,其特征是复杂的I缺陷,导致视网膜神经节细胞突然退化。尽管通常与线粒体DNA中的致病变异有关,但最近在携带核基因DNAJC30,NDUFS2和MCAT的双重变体的患者中描述了LHON。MCAT是线粒体脂肪酸合成(MTFA)的一部分,也是MECR,即线粒体反式2-烯酰基-COA还原酶。MECR突变导致肌张力障碍,视神经萎缩和基底神经节异常的隐性儿童期综合症。我们通过整个外显子组测序研究了两个姐妹,这些姐妹在年轻时会突然且无痛的视觉丧失影响,部分恢复和持续的中央Scotoma。我们对酵母中的候选变体进行了建模,并研究了酵母和成纤维细胞中的线粒体功能障碍。我们测试了蛋白质脂酰化和细胞对酵母中氧化应激的反应。结果两个姐妹在MECR中携带了纯合致病变体(P.Arg258Trp)。在酵母中,MeCr-R258W突变体显示出氧化的障碍,氧气消耗率降低了30%,蛋白质水平降低了80%,表明结构不稳定。成纤维细胞证实了MECR蛋白的量减少,但未能再现Oxphos缺陷。呼吸络合物组装正常。最后,酵母突变体缺乏关键代谢酶的脂酰化,并且对H 2 O 2治疗更敏感。脂肪酸补充部分挽救了生长缺陷。结论,我们报告了第一个具有纯合MECR变体的家庭,导致LHON样神经病,该杂志是最近的MCAT发现,增强了MTFAS作为LHON新型致病机制的损害。

计算模型用于识别针对以糖酵解增强为特征的转移性去势抵抗性前列腺癌的药物
摘要:由于缺乏有效的治疗方法,转移性去势抵抗性前列腺癌 (mCRPC) 仍然是一种致命疾病。癌症代谢向糖酵解升高方向重编程是 mCRPC 的标志。我们的目标是确定与高糖酵解特别相关的治疗方法。在这里,我们建立了一个计算框架,以在肿瘤微环境下识别具有增强糖酵解活性的 mCRPC 的新药物,然后进行体外验证。首先,使用我们已建立的计算工具 OncoPredict,我们估算了来自两个大型临床患者队列的每个 mCRPC 肿瘤中大约 1900 种药物对药物反应的可能性。我们选择了预测敏感性与糖酵解评分高度相关的药物。总共确定了 77 种预测在高糖酵解 mCRPC 肿瘤中更敏感的药物。这些药物代表了不同的作用机制。基于在泛癌细胞系中与糖酵解/OXPHOS 相关的最高测量药物反应,我们选择了三种候选药物伊维菌素、CNF2024 和 P276-00 进行后续体外验证。通过降低培养基中的输入葡萄糖水平以模拟 mCRPC 肿瘤微环境,我们在 PC3 细胞中诱导了高糖酵解条件,并验证了在此条件下这三种药物预计的更高敏感性(所有药物的 p < 0.0001)。对于生物标志物的发现,预测伊维菌素和 P276-00 对具有低雄激素受体活性和高糖酵解活性(AR(低)Gly(高))的 mCRPC 肿瘤更敏感。此外,我们整合了蛋白质-蛋白质相互作用网络和拓扑方法来识别这些候选药物的生物标志物。通过多个独立的生物标志物提名管道,EEF1B2 和 CCNA2 分别被确定为伊维菌素和 CNF2024 的关键生物标志物。总之,这项研究通过精准靶向高糖酵解的 mCRPC,提供了超越传统雄激素剥夺疗法的新型有效治疗方法。

线粒体的自然和人工机制...
在发现DNA双螺旋的十年后,Magrit和Sylvan Nass发现线粒体具有自己的双链DNA(线粒体DNA,mtDNA),其结构与核DNA的差异[1]。目前,已经确定,长度为16,659 bp的人类圆形mtDNA编码37个基因,这对于细胞氧化磷酸化(OXPHOS)和稳定的能量产生至关重要[2,3]。高度紧凑的mtDNA具有与核基因组不同的许多特征[4],其中一些特征导致mtDNA的独特遗传漂移。线粒体基因组的传播严格是母体。单亲遗传是在大鼠[5]和小鼠[6]中首次显示的,然后在人类中确认[7]。尽管有几项研究辩论了两种族裔mtDNA的遗传[8-11],但这种情况似乎非常罕见,并且它们的机制在很大程度上尚不清楚。其次,线粒体基因组以多个副本存在。一个SO-MATIC细胞含有约10 3 –10 4分子mtDNA [12,13]。最常见的是,一个细胞只有一种类型的线粒体基因组。这种情况 - hom质 - 通常会在mtDNA的所有等位基因膨胀时发生。但是,考虑到与核DNA相比的mtDNA突变率的增加,mtDNA的有缺陷拷贝可以与细胞中的野生型等位基因共存[14-17]。这种现象称为异质是线粒体基因组的特异性。多细胞生物中的不同细胞可能具有多种mtDNA物种。这种异质性可能在不同的发展阶段发生,并朝着不可限制的方向进行。更重要的是,异质的水平可能在一个人的器官之间以及一个单个家族中的个体之间在同一组织或器官的细胞之间变化[18]。定义异质质的原因,动力学和驱动力将提供有关许多线粒体疾病的遗传和进展的见解,在人群中,其流行率约为1:4300 [19]。

防御脂液滴:专为抗菌免疫设计的细胞器
fi g u r e 1脂质液滴:代谢,形态和组成。(a)主要代谢途径和中间代谢产物的简化方案参与LDS的生物发生和消耗。有关其他详细信息,请参见文本。fa,脂肪酸; FA-COA,酰基辅酶A; CPT1,肉碱棕榈转移酶I; CAC,柠檬酸周期; FASN,脂肪酸合酶; Oxphos,氧化磷酸化; ACC,乙酰辅酶A羧化酶; GPAT,甘油-3-磷酸酰基转移酶; AGPAT,1-酰基-SN-甘油-3-磷酸酰基转移酶; PAP,磷脂酸磷酸酶; DGAT,二甘油类酰基转移酶-1和-2; ACSL,酰基-COA合成酶; ATGL,脂肪甘油三酸酯脂肪酶; HSL,激素敏感脂肪酶; MAGL,单酰基甘油脂肪酶; NCEH,中性胆固醇酯水解酶。(b)内质网中发生的LD生物发生的示意图(ER)。酯化后,中性脂质积聚在ER双层中,形成透镜结构,该结构在ER双层内经过相位分离并成长为形成新生LD的细胞质。细胞质和ER蛋白被募集到LDS表面,促进其生长并萌芽到成熟的LDS中。附件蛋白在此过程中合作。在上面板(红色:TAG的化学结构)中说明了脂肪酸(FA)到三酰基甘油(TAG)中的酯化。(c)。用油酸处理肝HuH7细胞以诱导LD形成16小时(左图)。plin2(绿色)用特异性抗体定位,并用Lipidtox染色中性脂质。(n)表示细胞的核。箭头标记高放大倍数插图中的LD。THP-1细胞进行TEM分析(右图)。脂质液滴由它们的球形形态,相对较低的电子密度和通过单个磷脂单层界定。(d)代表LDS上主要蛋白质的简化方案。(e)该方案包含一些由病原体在宿主细胞中分泌的毒力因子操纵的LD蛋白(黑色)的例子(红色)(有关详细信息,请参见文本)。

肿瘤干细胞的多维分析
As a key factor in tumorigenesis, progression, recurrence and metastasis, the biological properties, metabolic adaptations and immune escape mechanisms of CSCs are the focus of current oncological research.CSCs possess self-renewal, multidirectional differentiation and tumorigenicity, and their mechanisms of action can be elucidated by the clonal evolution, hierarchical model and the dynamic CSCs model, of which the dynamic model is widely recognized due to its better explanation of the function and origin of CSCs.The origin hypothesis of CSCs involves cell-cell fusion, horizontal gene transfer, genomic instability and microenvironmental regulation, which together shape the diversity of CSCs.In terms of classi fi cation, CSCs include primary CSCs (pri-CSCs), precancerous stem cells (pre-CSCs), migratory CSCs (mig-CSCs), and chemo-radiotherapy-resistant CSCs (cr-CSCs and rr-CSCs), with each type playing a speci fi c role in tumor progression.Surface markers of CSCs, such as CD24, CD34, CD44, CD90, CD133, CD166, EpCAM, and LGR5, offer the possibility of identifying, isolating, and targeting CSCs, but the instability and heterogeneity of their expression increase the dif fi culty of treatment.CSCs have adapted to their survival needs through metabolic reprogramming, showing the ability to fl exibly switch between glycolysis and oxidative phosphorylation (OXPHOS), as well as adjustments to amino acid and lipid metabolism.The Warburg effect typi fi es their metabolic pro fi les, and altered glutamine and fatty acid metabolism further contributes to the rapid proliferation and survival of CSCs.CSC能够通过调节代谢网络来保持其干性特征,增强抗氧化剂防御并适应治疗应力来维持其干性。免疫逃生是CSC维持其生存的另一种策略,CSC可以通过诸如调节PD-L1表达的机制有效地逃避免疫监视,并促进免疫抑制性微环境的形成。一起,这些特性揭示了CSC的多维复杂性,强调了对CSC生物学对开发更有效肿瘤治疗策略的发展的重要性。将来,针对CSC的疗法将集中于表面标记物的精确鉴定,代谢途径的干预以及克服免疫逃生,以改善癌症治疗的相关性和效率,并最终改善患者的预后。
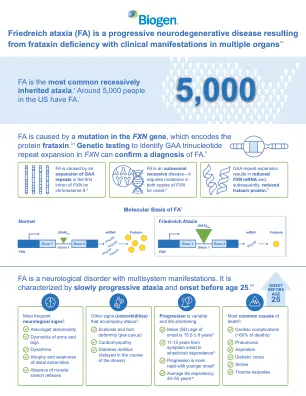
弗里德赖希共济失调 (FA) 是一种由 frataxin 缺乏引起的进行性神经退行性疾病,临床表现涉及多个器官 1
ARE,抗氧化反应元件;ATP,三磷酸腺苷;DNA,脱氧核糖核酸;FA,弗里德赖希共济失调;GAA,鸟嘌呤腺嘌呤腺嘌呤;ISC,铁硫簇;Keap1,Kelch 样 ECH 相关蛋白 1;Nrf2,核因子红细胞 2 相关因子 2;OXPHOS,氧化磷酸化;ROS,活性氧;SD,标准差。参考文献:1. 弗里德赖希共济失调研究联盟。什么是 FA?可从 https://www.curefa.org/understanding-fa/what-isfriedreichs-ataxia/ 获取。访问日期:2024 年 11 月。2. Koeppen AH。J Neurol Sci。2011;303(1-2):1-12。3. Campuzano V 等人。Hum Mol Genet。 1997;6(11):1771-1180。 4.Nachun D 等人。哈姆·摩尔·热内特。 2018;27(17):2965-2977。 5.弗里德赖希共济失调研究联盟。 Friedreich 共济失调临床管理指南 (FRDA)。可从 https://frdaguidelines.org/ 获取。访问时间:2024 年 11 月。 6. Campuzano V 等人。科学 。 1996;271(5254):1423-1427。 7.Gatchel JR 等人。纳特·热内特。 2005;6(10):743-755。 8. Bürk K. 小脑共济失调。 2017;4:4。 9.潘道夫·M·尼罗尔·吉内特。 2020;6(3):e415。 10. 汉森 E 等人。世界心脏病杂志。 2019;11(1):1-12。 11.Chiang S 等。神经化学国际公司。 2018;117:35-48。 12. González-Cabo P,帕劳 F. J Neurochem。 2013;126(补编1):53-64。 13. Llorens JV 等。神经科学前沿。 2019;13:75。 14. Petrillo S 等人。国际分子科学杂志。 2017;18(10):2173。 15.D'Oria V 等人。国际分子科学杂志。 2013;14(4):7853–7865。 16. Itoh K 等人,基因发育. 1999;13(1):76-86。17. Santos R 等人,抗氧化还原信号. 2010;13(5):651-690。

286疫苗增强的汽车T串扰宿主免疫...
背景嵌合抗原受体(CAR)T细胞疗法在实体瘤中令人失望。一个关键的障碍是先前存在的抗原异质性,并非所有肿瘤细胞表达汽车靶向的抗原。抗原分布(AS)是对与原始疗法靶标不同的二级抗原的免疫反应的诱导和扩增。可以在抗肿瘤周围T细胞库的扩张和扩展中发挥重要作用。我们最近开发了一种合成疫苗(AMPH-VAX),以通过直接调节CAR T细胞来增强对实体瘤的CAR T细胞活性,并通过AS 1 2通过AS 1 2(图1A)吸收宿主免疫。方法通过将car含量(即本提案中的pepviii)连接到白蛋白结合聚(乙二醇)磷脂来产生AMPH-VAX,然后用环状-DI-GMP(Sting Agonist)配制。我们使用了表达肿瘤特异性表面抗原EGFRVIII的CT-2A鼠胶质母细胞瘤模型。结果在CD4+和CD8+ T-Cell室中都引起了明显的抗原分散(AS),但不单独使用CAR T疗法Amph-Vax增强了CAR T(CAR T-VAX)治疗(图1B)。21,835个肿瘤内源性T细胞的单细胞RNA-SEQ证实了细胞毒性CD8 T细胞的显着增加和TH1 CD4 T细胞的诱导。跨性别分析表明,疫苗促进疫苗的增强显着增强了CAR T细胞代谢,包括氧化磷酸化(OXPHOS)。使用PGC-1 A-缺陷型CAR T-VAX治疗减少了〜50%。IFN G封锁被废除为(图1C,d)。IFN G封锁被废除为(图1C,d)。使用抗IL12(p75)抗体或IL12RB2基因敲除小鼠的IL12信号传导的封锁态ifn g封底,导致可忽略不计为(图1E)。我们进一步观察到在CD11C+ DC中IFN G信号不足的BATF3二氧化体小鼠或小鼠中的AS显着降低(图1F)。最后,我们表明IFNGR1-或IL12RB2缺陷型CAR T-VAX治疗未能诱导AS。使用EGFRVIII+和EGFRVIII-CT-2A细胞以预定比率混合的异质性肿瘤模型,我们发现CAR-T VAX治疗的〜50%动物携带的肿瘤含有高达20%EGFRVIII的肿瘤,并进一步升高了CAR T-IFN G表达的升高率增加了Cure Cure速率(增加了80%),并增加了80%(图)。和具有内源性T细胞且因此没有抗原扩散的RAG1 - / - 小鼠中的治疗反应完全损失(图1G)。结论是通过与DC衍生的IL-