XiaoMi-AI文件搜索系统
World File Search Systemacidic

合金钢在高酸性地热中的适用性...
本研究旨在评估目前市场上哪种材料可以克服腐蚀问题并承受火山环境中地热资源开采中的高腐蚀性条件。我们的调查是由 Lahendong 地热田(印度尼西亚北苏拉威西岛)的条件引发的:LHD-23 井是最大的挑战之一,因为它能够从单个井产生 > 20 MW 的能量,同时具有非常低的 pH 值(2 - 3)和相对较高的氯化物(1,500 mg/L)和硫酸盐(1,600 mg/L)浓度。选择了三种不同的钢种(低合金钢 UNS G41300、不锈钢 UNS S31603 和高合金不锈钢 UNS N08031),并通过短期电化学方法(动电位极化)和长期暴露试验(最长 6 个月)评估了它们的腐蚀行为。该研究在实验室中在 100°C(100 kPa)和 175°C(900 kPa)的人工 LHD-23 地热盐水(1,500 mg/L 氯化物、1,600 mg/L 硫酸盐、pH 2)的静止条件下进行,模拟现场的条件。
合金钢在高酸性地热中的适用性...
本研究旨在评估目前市场上哪种材料可以克服腐蚀问题并承受火山环境中地热资源开采中的高腐蚀性条件。我们的调查是由 Lahendong 地热田(印度尼西亚北苏拉威西岛)的条件引发的:LHD-23 井是最大的挑战之一,因为它能够从单个井产生 > 20 MW 的能量,同时具有非常低的 pH 值(2 - 3)和相对较高的氯化物(1,500 mg/L)和硫酸盐(1,600 mg/L)浓度。选择了三种不同的钢种(低合金钢 UNS G41300、不锈钢 UNS S31603 和高合金不锈钢 UNS N08031),并通过短期电化学方法(动电位极化)和长期暴露试验(最长 6 个月)评估了它们的腐蚀行为。该研究在实验室中在 100°C(100 kPa)和 175°C(900 kPa)的人工 LHD-23 地热盐水(1,500 mg/L 氯化物、1,600 mg/L 硫酸盐、pH 2)的静止条件下进行,模拟现场的条件。

simoa®神经纤维纤维酸性蛋白(GFAP)
神经系统疾病仍然是早期诊断和治疗最具挑战性的疾病之一。与“可见”疾病不同,神经元损伤和神经变性可能会被忽视或误认为其他情况。症状的微妙和当今评估的主观性质也使得很难尽早确定这些疾病。当前,对于早期检测神经退行性疾病(例如AD),尚无确切的测试。临床医生只能在症状开始出现后才最终诊断它们。因此,许多患者可能会等待数年的诊断或明确的治疗途径。
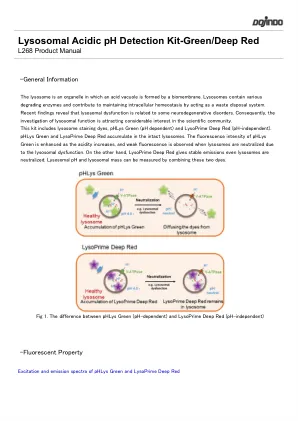
溶酶体酸性pH检测试剂盒绿色/深红色
最近的发现表明,溶酶体功能障碍与某些神经退行性疾病有关。因此,对溶酶体功能的调查引起了科学界的极大兴趣。该套件包括溶酶体染色染料,phlys绿色(pH依赖)和溶液深红色(无pH无依赖性)。

恶唑:在酸性环境中腐蚀抑制钢的有前途的边界
酸性环境中的钢腐蚀是经济各个部门的严重问题。必须通过制定有效的腐蚀保护计划来控制它。在这篇综述中,总结了酸溶液中的铁衍生物作为铁抑制剂。首先描述酸性腐蚀和钢抑制钢作为控制腐蚀的手段的腐蚀。 然后引入了恶唑衍生物作为腐蚀抑制剂的可能性。 详细介绍了沙唑衍生物可以防止金属腐蚀的均值。 描述了经典方法和合成新的恶唑的最新趋势,尤其是获得恶唑衍生物的旅程。 侧重于奥沙唑的抑制作用,影响其效率的因素以及与其他抑制作用的比较分析的实验室研究。 也讨论了氧化唑作为石油和天然气,化学加工,汽车,海洋水处理行业的腐蚀抑制剂的工业应用。 对未来研究的关注和可能性以及如何利用奥卡唑来耐腐蚀,将扩大我们对科学界腐蚀的了解。 这项研究证明了恶唑作为腐蚀抑制剂及其重要性的潜力。 它为改善酸性环境中钢的腐蚀处理提供了新的想法。酸性腐蚀和钢抑制钢作为控制腐蚀的手段的腐蚀。然后引入了恶唑衍生物作为腐蚀抑制剂的可能性。详细介绍了沙唑衍生物可以防止金属腐蚀的均值。经典方法和合成新的恶唑的最新趋势,尤其是获得恶唑衍生物的旅程。侧重于奥沙唑的抑制作用,影响其效率的因素以及与其他抑制作用的比较分析的实验室研究。也讨论了氧化唑作为石油和天然气,化学加工,汽车,海洋水处理行业的腐蚀抑制剂的工业应用。对未来研究的关注和可能性以及如何利用奥卡唑来耐腐蚀,将扩大我们对科学界腐蚀的了解。这项研究证明了恶唑作为腐蚀抑制剂及其重要性的潜力。它为改善酸性环境中钢的腐蚀处理提供了新的想法。

变性剂可以稳定DNA吗?吡啶逆转酸性pH **
DNA的生物学作用与水溶液中的结构和稳定性密切相关。DNA的完全脱水或用较低极性的溶剂取代水会导致DNA结构发生很大变化[1]。溶液的离子强度的变化产生了显着的结构可塑性[2],并且柜台性质的变化甚至可以逆转双链DNA稳定性的规范规则[3]。 某些渗透液,例如尿素,甲酰胺,氯化硫苷,二甲基硫氧化物或吡啶是化学变性剂[4]。 我们最近使用了微秒长的分子动力学(MD)模拟来证明PYR的非常强的变性特性与其通过在开放的,溶剂暴露的核碱基上堆叠捕获显微镜展开事件的能力有关[5]。 在这里,我们在另一种强大的变性剂的存在下探索了PYR的变性特性:pH。 在这项工作中,我们评估了这两种变性剂的效果是加性,合作或抗合作性的。 我们首先探索了具有不同GC含量的三种DNA双链体在中性pH值下的分泌特性(表1)。 图1(a,b)和补充表S1 所示的结果溶液的离子强度的变化产生了显着的结构可塑性[2],并且柜台性质的变化甚至可以逆转双链DNA稳定性的规范规则[3]。某些渗透液,例如尿素,甲酰胺,氯化硫苷,二甲基硫氧化物或吡啶是化学变性剂[4]。我们最近使用了微秒长的分子动力学(MD)模拟来证明PYR的非常强的变性特性与其通过在开放的,溶剂暴露的核碱基上堆叠捕获显微镜展开事件的能力有关[5]。在这里,我们在另一种强大的变性剂的存在下探索了PYR的变性特性:pH。在这项工作中,我们评估了这两种变性剂的效果是加性,合作或抗合作性的。我们首先探索了具有不同GC含量的三种DNA双链体在中性pH值下的分泌特性(表1)。图1(a,b)和补充表S1

将大麦谷物提取物作为碳钢
从不同温度下,大麦种子提取物对1M盐酸在1M盐酸中腐蚀的作用是从它们作为绿色抑制剂在清洁和降水过程中的潜在用途的角度研究的。使用50%乙醇/水(VOL)溶液进行提取。使用了两种提取方法:浸泡和超声。通过通过电化学方法(Potentiodynalicallization(Tafel曲线)和电化学耐药性光谱)研究吸附和腐蚀过程来研究抑制剂的抑制作用机制。研究结果没有表明提取方法对抑制行为没有影响,抑制作用作为浓度的函数显示,抑制效率的抑制效率显着提高到浓度为400 ppm,然后在这两种方法中都与其无关。然而,浸泡方法的抑制效率在800 ppm时(87.01%,而超声方法为80%)。对该提取物的抑制机制的研究表明化学吸附的可能性。抑制活性随温度增加。抑制活性随温度增加。

在三唑衍生物的计算机研究中,作为酸性培养基中低碳钢的腐蚀抑制剂
理论方法(例如量子计算和Monte Carlo(MC)模拟,由于学习结构的相对较快方法,在研究腐蚀抑制剂方面非常重要。在本文中,利用了几种半经验量子计算方法(AM1,PM3和PM6)来研究某些三唑的腐蚀抑制效率(CIE),作为金属腐蚀的抑制剂(降低了降低了降低了至1 m盐酸)。MC模拟技术用于本研究来计算吸附能。优化的基态几何形状,最高占用分子轨道(E HOMO)的能级,最低无占用分子轨道(E Lumo)的能级,吸附能和偶极矩(μ)与三氮化衍生物的CIE相关。建议三个方程式计算CIE。在CIE EXP和CIE计算之间发现了良好的协议。CIE EXP和CIE CALC之间的相关系数(R)位于0.931至0.955之间。AM1,PM3和PM6可有效测量CIE。 回归分析在非线性方程中包含吸附能时使用的量子参数较少。 e广告可以减少描述符数量,以创建易于使用和短暂的模型。AM1,PM3和PM6可有效测量CIE。回归分析在非线性方程中包含吸附能时使用的量子参数较少。e广告可以减少描述符数量,以创建易于使用和短暂的模型。

Lepidium didymium植物提取物作为酸性培养基中的钢的环保腐蚀抑制剂
通过在1 M H 2 SO 4中应用体重减轻分析,SEM和光谱技术,在碳钢腐蚀上应用体重减轻分析,SEM和光谱技术来探索鳞翅目didymiu M的空中抑制效率。这些技术已被用来研究钢的腐蚀特征,在不存在和存在各种量的鳞翅目Didymiu M提取物的情况下。借助减肥统计,研究了植物提取物在钢表面上构建防御膜的能力。表面形态分析(SEM)也支持了提取物在钢表面上的保护膜的形成。在2500 mg/l时,植物提取物在1 m H 2中具有最佳的钢抑制效率,因此4是91.16%。鳞霉菌提取物被认为是有效的抑制剂。

可可豆壳作为酸性培养基抑制剂的电化学和经济评估
通过电化学技术和重量测量结果研究了可可豆壳粉(CBSP)对SAE 1008碳钢的抑制作用。傅立叶变换红外光谱(FTIR)和扫描电子显微镜(SEM)用于化学和形态表征。重量表和EIS结果表明,腐蚀速率随抑制剂浓度的增加而降低,在1.77 g L -1抑制剂中达到97.92%的最大值(最高浓度研究)。即使在最低浓度(0.44 g l -1)也观察到良好的腐蚀抑制效率(96.03%)。极化曲线表明CBSP充当混合类型抑制剂。抑制性分子吸附遵循Langmuir等温模型。抑制剂经济评估表明,CBSP的使用比传统抑制剂便宜48-616倍。结果表明,CBSP是碳钢的有效腐蚀抑制剂,其生产在财务上具有吸引力。