XiaoMi-AI文件搜索系统
World File Search Systemdpt
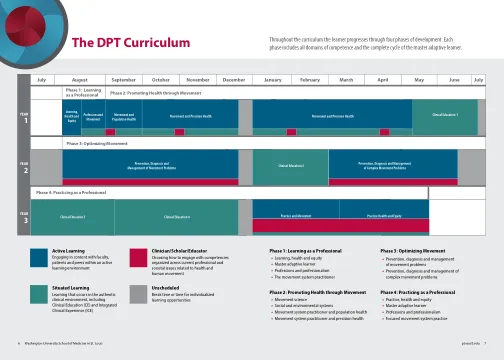

OTD 和 DPT COVID 疫苗接种程序
值得注意的是,即使医疗机构允许豁免,多个地区组织也表示,由于相关风险,他们不会接受未接种疫苗的学生。在与 DPT 顾问委员会成员的交谈中,有一些案例表明,接种过疫苗的治疗师不想与未接种疫苗的人一起工作,患者拒绝与未接种疫苗的治疗师一起工作,甚至有组织将疫苗接种状况纳入急性和长期护理机构聘请治疗师的申请流程中。当组织允许豁免时,一些机构会要求学生每周至少接受一次 COVID-19 检测和/或佩戴特殊的个人防护设备。
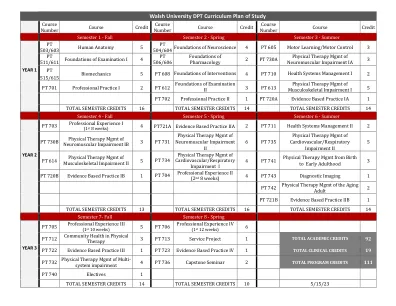

Katrina低啤酒,PT,DPT,ATP 609-903-2030 合作伙伴学校课程等效
参加了许多主题的讲座,包括:研究;治疗;伦理;等。2020年2月,丹佛,Co Dahl C,Lawton C,Low-Beer K,Pol Lak M,SmithS。跑步机培训对运动功能,功能性移动性和唐氏综合症婴儿生活质量的影响。论文发表于:克拉克森大学; 2015年12月;纽约州波茨坦。低啤酒K.在patie nt中加强,协调,步态和平衡训练,其中有潜在的小脑共济失调S/P下半部血肿和s骨/coccyx骨折:病例报告。论文发表于:克拉克森大学; 2015年9月;纽约州波茨坦。

估计DPT计划中的间接入学成本
个人笔记本电脑计算机(要求;估算):$ 1,200。计算机的价格差异很大,取决于模型和配置。大学笔记本电脑需求教科书(估算):〜$ 3,000(〜$ 400/学期)。教科书可以通过出版商网站或卫理公会大学书店购买。解剖软件:65美元。在程序期间使用的软件将下载到个人计算机上。课程供应和运输费用 - 个人课程可能需要额外的分配费用和参与社区活动。这些可能包括但不限于用于作业和演示的海报和小册子的印刷费用,前往社区网站的汽油,购买课堂项目的用品等。将在每个课程开始时或在作业/演示/活动之前提前通知这些费用中的任何一项。PT技能设备袋(必需):成本各不相同,约100美元,具体取决于设备的选择。磨砂膏(强烈建议用于解剖实验室):$ 50(变化)

Alison Lynn McKenzie,PT,MA,DPT,Ph.D。 战略计划 双年展评论:学年2020-2021&... 居民董事会和通勤进餐计划请求
Alison Lynn McKenzie,PT,M.A.,DPT,Ph.D。 解剖学实验室运营部物理治疗系物理治疗教授Crean健康与生命科学学院Harry和Diane Rinker Campus Chapman University 9401 Jeronimo Rd。 Irvine,CA 92618(714)-744-7827;手机:949-680-5230电子邮件:amckenzi@chapman.edu; almckenzi@gmail.com教育:Chapman University,CA 2008-2012物理治疗博士(DPT)过渡性物理治疗医生计划,2012年加利福尼亚大学伯克利分校,伯克利大学和旧金山伯克利,加利福尼亚州和旧金山,加利福尼亚州和旧金山,加利福尼亚州,加利福尼亚州,1990年至1991年1990年至1991年1991年,在加利福尼亚州伯克利亚郡的CA CAROELEEL,CA CARIMY pHEELEELIAT PHERKEELEELIAT PHERKEELEELYINY CARKEELEELYYENICEELEY,在解剖学,论文:丰富的环境中:研究康复对运动皮质病变后神经恢复的影响的模型。 加利福尼亚大学伯克利分校,加利福尼亚州伯克利分校,1987年硕士 在解剖学论文中:富集环境对906天大的长evans大鼠运动皮层的影响。 犹他大学盐湖城,犹他州研究生院先决条件课程,1982- 1983年,犹他州盐湖城,UT 1977-81 B.S. 物理治疗(Magna cum La)许可信息/注册号:Alison Lynn McKenzie,PT,M.A.,DPT,Ph.D。解剖学实验室运营部物理治疗系物理治疗教授Crean健康与生命科学学院Harry和Diane Rinker Campus Chapman University 9401 Jeronimo Rd。Irvine,CA 92618(714)-744-7827;手机:949-680-5230电子邮件:amckenzi@chapman.edu; almckenzi@gmail.com教育:Chapman University,CA 2008-2012物理治疗博士(DPT)过渡性物理治疗医生计划,2012年加利福尼亚大学伯克利分校,伯克利大学和旧金山伯克利,加利福尼亚州和旧金山,加利福尼亚州和旧金山,加利福尼亚州,加利福尼亚州,1990年至1991年1990年至1991年1991年,在加利福尼亚州伯克利亚郡的CA CAROELEEL,CA CARIMY pHEELEELIAT PHERKEELEELIAT PHERKEELEELYINY CARKEELEELYYENICEELEY,在解剖学,论文:丰富的环境中:研究康复对运动皮质病变后神经恢复的影响的模型。 加利福尼亚大学伯克利分校,加利福尼亚州伯克利分校,1987年硕士 在解剖学论文中:富集环境对906天大的长evans大鼠运动皮层的影响。 犹他大学盐湖城,犹他州研究生院先决条件课程,1982- 1983年,犹他州盐湖城,UT 1977-81 B.S. 物理治疗(Magna cum La)许可信息/注册号:Irvine,CA 92618(714)-744-7827;手机:949-680-5230电子邮件:amckenzi@chapman.edu; almckenzi@gmail.com教育:Chapman University,CA 2008-2012物理治疗博士(DPT)过渡性物理治疗医生计划,2012年加利福尼亚大学伯克利分校,伯克利大学和旧金山伯克利,加利福尼亚州和旧金山,加利福尼亚州和旧金山,加利福尼亚州,加利福尼亚州,1990年至1991年1990年至1991年1991年,在加利福尼亚州伯克利亚郡的CA CAROELEEL,CA CARIMY pHEELEELIAT PHERKEELEELIAT PHERKEELEELYINY CARKEELEELYYENICEELEY,在解剖学,论文:丰富的环境中:研究康复对运动皮质病变后神经恢复的影响的模型。加利福尼亚大学伯克利分校,加利福尼亚州伯克利分校,1987年硕士在解剖学论文中:富集环境对906天大的长evans大鼠运动皮层的影响。犹他大学盐湖城,犹他州研究生院先决条件课程,1982- 1983年,犹他州盐湖城,UT 1977-81 B.S.物理治疗(Magna cum La)许可信息/注册号:
伊朗2~6月龄婴儿五联疫苗与百白破疫苗不良事件比较:全国性研究
背景:疫苗接种是公共卫生中最显著的干预措施,是控制婴儿传染病的有效策略。目的:本研究旨在比较伊朗 2 至 6 个月大婴儿接种五联疫苗和 DPT 疫苗的不良事件。方法:这是一项分析性横断面研究,研究了 2013 年接种 DPT 疫苗和 2015 年接种五联疫苗的 2 至 6 个月大的健康婴儿是否经历过与这两种疫苗相关的不良事件。使用百分比、平均值、标准差和卡方检验来描述和分析数据(p < 0.05)。结果:结果显示,在伊朗各地接种这两种疫苗的婴儿中,分别记录了 10,464 起和 17,561 起与 DPT 疫苗和五联疫苗相关的不良事件。马赞德兰省、加兹温省和戈勒斯坦省报告的不良事件数量最多(分别为 15.74%、11.25% 和 9.12%)。此外,五联疫苗似乎比 DPT 有更多记录的不良事件,DPT 疫苗的高烧记录率最高(47.4%),五联疫苗的轻度局部并发症最高(31.68%)。疫苗种类与反应类型、不良事件分类和生产疫苗的国家之间存在显著关系(p < 0.05)。结论:与 DPT 疫苗相比,五联疫苗发生的高烧、呕吐、腹泻和烦躁等严重局部不良事件似乎较少。因此,在婴儿中用五联疫苗代替 DPT 疫苗似乎可以减少其中的不良事件。

Alison Lynn McKenzie,PT,MA,DPT,Ph.D。
我们继续做出我们的承诺,以改善查普曼体验的各个方面,以更好地为我们的学生,员工,教职员工和朋友服务。我们为塑造了这一计划的各种观点感到自豪,并受到已经实现的结果的启发,从新的学生住房到建筑物和福勒工程学院的启动。这些努力将不仅有助于查普曼大学,而且几十年来。我们专注于建立包容性社区和优化我们的校园足迹正在如火如荼地进行。我们致力于将查普曼提升到国家舞台,这是一个重视研究和创造性活动的机构。通过我们的综合努力,我们认为我们的成就只会继续增长。让我们庆祝战略计划每个领域中发生的一些里程碑。

BMVg 合约价目表
远程部分至 + 和 - 6.0 dpt 圆柱。对于 > +/-6 dpt 和 = < +8/-10 dpt 高折射率玻璃球面,最多收取 + 4.0 dpt 附加费到 +8 dpt 或至 - 11 dpt,圆柱。高达 +4 dpt,折射率高达 1.67 不可与数字 290 组合,订单上必须有德国联邦国防军处方医生的注释
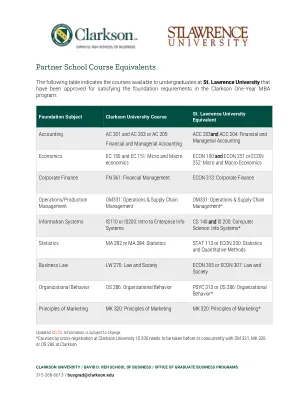
Katrina低啤酒,PT,DPT,ATP 609-903-2030合作伙伴学校课程等效
合作伙伴学校课程等效于下表指示圣劳伦斯大学大学生可用的课程,这些课程已获得批准满足克拉克森一年MBA课程的基础要求: