XiaoMi-AI文件搜索系统
World File Search SystemdsDNA

利用CRISPR/Cas9基因编辑技术治疗β-地中海贫血的最新进展
dsDNA 或 ssODN 作为模板进行精确修复 , 而非同源末端连接 (NHEJ) 介导的随机修复可造成插入 、 缺失或突变 . ssODN: 单链寡核苷酸 ; dsDNA: 双链 DNA Figure 3 Two CRISPR/Cas9 gene editing strategies. Cas9 creates DNA double strand break at three bases upstream of the PAM sequence. Homologous recombination repair (HDR) mediates precise repair using dsDNA or ssODN as a template, while non-homologous end joining (NHEJ) -mediated repair can cause insertion, deletion or mutation. ssODN: Single-strand oligodeoxynucleotide; dsDNA: Double strand DNA

测量蛋白质-DNA 凝聚物中的桥接力
蛋白质-DNA 凝聚物介导转录并调节基因表达以及 DNA 复制和修复。稳定凝聚物的分子间桥接力在这些过程中起着直接作用。在这里,我们使用光镊来测量桥接力。在鱼精蛋白存在的情况下,在两个微珠之间连接的 20.5 knt 单链 DNA (ssDNA) 上观察到单个凝聚物。拉伸产生具有锯齿状图案的力曲线,表明凝聚物是通过单个鱼精蛋白-ssDNA 桥的连续断裂而分解的。桥接力为 11.3 ± 4.6 pN,单个桥的展开长度为 1.3 ± 0.8 µm。相反,双链 DNA (dsDNA) 形成鱼精蛋白桥接缠结,可以承受足够高的力 (~55 pN) 以分离链。 ssDNA 通过在回缩时过度拉伸种子缠结形成,在 dsDNA 的缺口处追踪未剥离的部分,但初始凝聚物具有足够的 ssDNA 与 dsDNA 比率以呈现液体状,如随后拉伸中的锯齿状图案所示。dsDNA 的存在将桥接力提高到 34 ± 8 pN,在添加外部 ssDNA 后恢复到 ~10 pN。根据这些单分子结果,鱼精蛋白-dsDNA 混合物形成固体状聚集体,需要添加 ssDNA 才能变成液滴。相反,添加 dsDNA 会减慢鱼精蛋白-ssDNA 液滴的融合。这项工作展示了桥接力的首次测量,并表明 ssDNA 与 dsDNA 比率可以调整蛋白质-DNA 凝聚物中桥接力的大小。

破译 DNA 的序列依赖性结构和...
图 1:双链 DNA 结构和构象灵活性的参数化和描述。(A)双链 DNA 分子的结构通过刚体变换参数化,其中轴系统跨越每个碱基对步骤 (ri)。这些参数描述了将碱基对步骤的轴系统映射到相邻轴系统的旋转和平移函数。(B)标准化流模型从正态分布的潜在空间映射到描述双链 DNA 结构的参数。映射是通过多层实现的,并且是可逆的。

双链DNA IgG抗体
局限性:对双链DNA(DSDNA)的IgG抗体的测量是半定量的。不应依靠这些抗体的反应性略有变化来预测全身性红斑狼疮患者(SLE)患者的临床变化。SLE患者疾病的临床耀斑可能不伴有DSDNA抗体反应性的变化。因此,仅抗DSDNA抗体结果就不足以指导疾病管理。

M.Manickavasagam博士...
酸研究,其能够裂解单链DNA(ssDNA)和RNA的能力。该酶源自曲霉细菌,对单链核酸表现出很高的特异性,同时对双链DNA(dsDNA)相对无损(DSDNA)。

PNA辅助dnazymes裂解双链DNA,用于具有高序列保真度的基因工程
摘要:Dnazymes已被广泛用于许多传感和成像应用中,但是自1994年发现以来,很少使用基因工程,因为它们的底物范围主要限于单链DNA或RNA,而遗传信息则存储在双链DNA(DSDNA)中。为了克服这一主要局限性,我们在这里报告了肽核酸(PNA)辅助双链DNA通过dnazymes(Panda)辅助的DNA迹象,这是将Dnazyme活性扩展到DSDNA的第一个例子。我们表明,熊猫在有效划痕或导致靶dsDNA上有双链破裂是可以编程的,靶DsDNA模仿了蛋白质核酸酶,并且可以充当分子克隆中的限制酶。除了比蛋白质酶小得多,在我们测试的条件下,熊猫还具有更高的序列保真度,这证明了其作为基因工程和其他生化应用的新型替代工具的潜力。

引用:Anderson EW,Fishbein J,Hong J等。喹啉酸是一种kynurenine/色氨酸途径代谢产物,与认知受损的TES div>相关联
抽象的客观抗DSDNA抗体(抗DSDNA)是SLE中所有分类方案的组成部分,并包括经过验证的活动指标中的一个域之一。抗DSDNA经常通过酶免疫测定(EIA)或crithidia luciliae免疫荧光试验(CLIFT)进行商业测量。通过通过两种不同的测定法测量这些抗体的临床影响,这项研究利用了良好的多种族/种族组合。方法所有患者符合至少一种经过验证的方案的SLE分类标准:美国风湿病学院,系统性红斑狼疮国际合作诊所和/或美国风湿病学/欧洲联盟反对风湿主义分类标准。通过多重EIA和Clift同时配对的抗DSDNA患者。 对一致性或不一致的分析,测定法的滴度可比性以及与杂交SLE病活动指数评分,狼疮肾炎的患病率(LN),预测耀斑和分类标准的能力。 结果207名患者由EIA和Clift至少一次用于抗DSDNA,产生了586个配对结果。 377对是一致的,有209对不一致。 207例患者中的41个总是不一致的成对结果,39名患者总是患有滴度不一致的结果。 在100例LN患者中,60例EIA为阳性,Clift为72例。 对LN与没有LN患者的患者的敏感性和特异性分别为EIA 60%和47%,Clift分别为72%和37%。通过多重EIA和Clift同时配对的抗DSDNA患者。对一致性或不一致的分析,测定法的滴度可比性以及与杂交SLE病活动指数评分,狼疮肾炎的患病率(LN),预测耀斑和分类标准的能力。结果207名患者由EIA和Clift至少一次用于抗DSDNA,产生了586个配对结果。377对是一致的,有209对不一致。207例患者中的41个总是不一致的成对结果,39名患者总是患有滴度不一致的结果。在100例LN患者中,60例EIA为阳性,Clift为72例。对LN与没有LN患者的患者的敏感性和特异性分别为EIA 60%和47%,Clift分别为72%和37%。42例成对结果后的90天内进行了耀斑评估。 七个轻度耀斑和四名患有严重耀斑的患者中有六名均具有一致的阳性结果。 结论我们的数据表明,抗DSDNA的两种测定法之间的阳性不一致相对普遍,发生在整个患者的五分之一和三分之一的访问中。 eiA阳性与LN相比,LN的频率少于Clift阳性。 与抗DSDNA分析之间的结果显着不一致,获得Clift和EIA分析可能对SLE的分类和常规监测可能是有益的。42例成对结果后的90天内进行了耀斑评估。七个轻度耀斑和四名患有严重耀斑的患者中有六名均具有一致的阳性结果。结论我们的数据表明,抗DSDNA的两种测定法之间的阳性不一致相对普遍,发生在整个患者的五分之一和三分之一的访问中。eiA阳性与LN相比,LN的频率少于Clift阳性。与抗DSDNA分析之间的结果显着不一致,获得Clift和EIA分析可能对SLE的分类和常规监测可能是有益的。
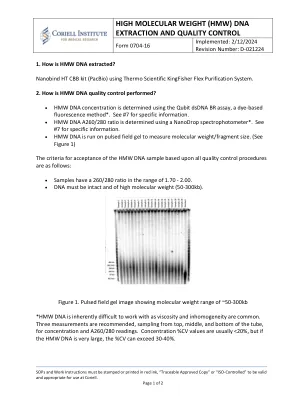
高分子量(HMW)DNA提取和质量控制
我们建议使用Qubit®DSDNABR分析(Q32850; Thermofisher Scientific)确定HMW DNA浓度。该测定法使用超敏感的荧光核酸染色来用标准的荧光计和荧光素激发和发射波长来量化双链DNA(dsDNA)。建议从顶部,中部和底部进行多次测量。

抗DSDNA抗体之间的临床意义是多种流量免疫测定和crithidia luciliae分析中的多种种族种族群体
1 Dobin,A。等。 星:超快通用RNA-seq对准器。 生物信息学29,15-21,doi:10.1093/bioinformatics/bts635(2013)。 2 Liao,Y.,Smyth,G。K.&Shi,W。R package rsubread更容易,更快,更便宜,更适合对RNA测序读取的对齐和定量。 核酸Res 47,E47,DOI:10.1093/nar/gkz114(2019)。 3 Love,M。I.,Huber,W。&Anders,S。带有DESEQ2的RNA-Seq数据的折叠变化和分散的调节估计。 基因组生物学15,550,doi:10.1186/s13059-014-0550-8(2014)。 4 Subramanian,A。等。 基因集富集分析:一种基于知识的方法,用于解释全基因组表达谱。 Proc Natl Acad Sci U S 102,15545-15550,doi:10.1073/pnas.0506580102(2005)。 5 Kolberg,L.,Raudvere,U.,Kuzmin,I.,Vilo,J. &Peterson,H。Gprofiler2-用于基因列表功能富集分析和名称空间转换工具集的R软件包G:Profiler。 f1000res 9,doi:10.12688/f1000research.24956.2(2020)。 6 Zhang,Y.,Parmigiani,G。&Johnson,W。E.战斗seq:RNA-Seq计数数据的批处理效应调整。 nar Genom Bioinform 2,LQAA078,doi:10.1093/nargab/lqaa078(2020)。1 Dobin,A。等。星:超快通用RNA-seq对准器。生物信息学29,15-21,doi:10.1093/bioinformatics/bts635(2013)。2 Liao,Y.,Smyth,G。K.&Shi,W。R package rsubread更容易,更快,更便宜,更适合对RNA测序读取的对齐和定量。核酸Res 47,E47,DOI:10.1093/nar/gkz114(2019)。3 Love,M。I.,Huber,W。&Anders,S。带有DESEQ2的RNA-Seq数据的折叠变化和分散的调节估计。基因组生物学15,550,doi:10.1186/s13059-014-0550-8(2014)。4 Subramanian,A。等。基因集富集分析:一种基于知识的方法,用于解释全基因组表达谱。Proc Natl Acad Sci U S 102,15545-15550,doi:10.1073/pnas.0506580102(2005)。5 Kolberg,L.,Raudvere,U.,Kuzmin,I.,Vilo,J.&Peterson,H。Gprofiler2-用于基因列表功能富集分析和名称空间转换工具集的R软件包G:Profiler。f1000res 9,doi:10.12688/f1000research.24956.2(2020)。6 Zhang,Y.,Parmigiani,G。&Johnson,W。E.战斗seq:RNA-Seq计数数据的批处理效应调整。nar Genom Bioinform 2,LQAA078,doi:10.1093/nargab/lqaa078(2020)。

海报:使用单链 DNA 模板进行高效的同源定向修复
(ssDNA) 是优于 dsDNA 的 HDR 模板。在此,我们报告了一项系统研究,比较了 HEK293 细胞中的 dsDNA HDR 模板和 IDT Ultramer® 寡核苷酸 ssDNA HDR 模板。测试了链选择和同源臂长度,以确定 HDR 在 Cas9 dsDNA 断裂点创建新的限制位点(6 个碱基插入)的效率。使用较长 ssDNA HDR 模板的初步实验表明,与较短 ssDNA 模板具有类似的优势和行为。具有不对称同源臂的模板的 HDR 插入。使用具有不对称同源臂的 HDR 模板导致 EcoRI 插入率与对称同源臂的插入率相似。使用靶向链模板获得了高 HDR 效率,其中