XiaoMi-AI文件搜索系统
World File Search SystemdsDNA
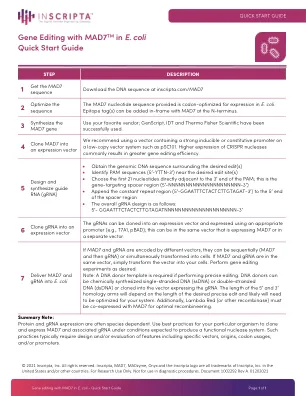
使用 MAD7TM 在大肠杆菌中进行基因编辑的快速入门指南
如果 MAD7 和 gRNA 由不同的载体编码,则可以依次(MAD7 然后是 gRNA)或同时将其转化为细胞。如果 MAD7 和 gRNA 在同一个载体中,只需将载体转化为细胞即可。根据需要进行基因编辑实验。注意:如果进行精确编辑,则需要 DNA 供体模板。DNA 供体可以是化学合成的单链 DNA (ssDNA) 或双链 DNA (dsDNA),也可以克隆到表达 gRNA 的载体中。5' 和 3' 同源臂的长度取决于所需精确编辑的长度,可能需要针对您的系统进行优化。此外,Lambda Red(或其他重组酶)必须与 MAD7 共同表达才能实现最佳重组。

sacituzumab govitecan在表达Trop-2的三阴性乳腺癌
sacituzumab govitecan(SG)是一种抗体 - 药物结合物,由通过水解链接器与SN-38的可胞样蛋白酶体I抑制性活性成分SN-38结合的人源化抗Trop-2 IgG抗体组成。我们研究了SN-38介导的双链DNA(DSDNA)的Trop-2表达和同源重组修复(HRR)是否在三阴性乳腺癌(TNBC)对SG的敏感性中起作用。在RAD51表达中所证明的HRR途径的激活在SG敏感的细胞系中评估了低和中度的Trop-2-表达(SK-MES-1鳞状细胞肺癌和HCC1806 TNBC),与低TROP-2-表达TROP-2-表达SG-SG-SESTENS敏感TNBC细胞系(MDA)相比。此外,在SG的小鼠中处理了MDA-MB-231,C13和C39的两种TROP-2-2-转移剂(分别为4和25倍)(分别为4和25倍),以确定Trop-2表达的增加是否提高了SG的效率。sg介导的MDA-MB-231中RAD51的增加> 2倍,但在SK-MES-1或HCC1806中没有影响,导致MDA-MB-231中DSDNA断裂水平较低。sg和盐水对父母MDA-MB-231肿瘤小鼠产生了相似的作用(中位生存时间(MST)= 21d和19.5d)。然而,在转染后具有较高trop-2表达C13和C39肿瘤的小鼠中,SG提供了显着的生存益处,即使与Irinotecan相比(MST = 97d vs. 35d vs. 35d vs. 35d,C13和81d vs.81d vs. 28d vs.28d vs. p <0.0007; p <0.0007; p <0.0007); p <0.0007)。这些结果表明,对于表达高水平Trop-2的HRR肿瘤患者以及对HRR缺陷型肿瘤表现出低/中等水平的Trop-2的患者,SG可以比Irinotecan提供更好的临床益处。

CRISPR 敲入协议 - 用于刺激人类 T 细胞......
注意:确保将 Cas9 蛋白作为反应中的最后一种材料添加。如果进行了敲除编辑,则无需添加 HDRT。表中 Cas9 与 sgRNA 的比例为 1:3。强烈建议对每个设计进行 Cas9:sgRNA 比例以及 Cas9 蛋白和 HDRT 的量进行实验优化。GenScript 建议通过测试 1:1 和 1:4 之间的 RNP 比例开始优化。ssDNA 或 dsDNA 的量可以分别最初设置在 1 μ g - 4 μ g 或 0.2 μ g - 1 μ g 之间(对于 0.5 kb 和 4 kb 之间的 HDRT;如果 HDRT 长度超出此范围,则需要相应调整试剂量)。最好为第一次实验设置阴性对照、阳性对照和转染对照。

HAMILTON NGS STARlet 上的 Oxford Nanopore 连接测序试剂盒 XL V14 自动化为 Nanopore Sequen 生成高质量 DNA 文库
资格设置和结果 为了在 NGS STARlet 上对 Oxford Nanopore SQK-LSK114-XL V14 V1.0 方法进行生物学验证,对 8 个(4 个阳性样本 + 4 个阴性对照)或 24 个样本(22 个阳性样本 + 2 个阴性对照)进行了生物学运行。作为输入材料,1 μg 全长(48 kB)噬菌体 Lambda DNA 用于 8 个样本的运行。对于 24 个样本的运行,1 μg 剪切(9kB)人类基因组 DNA 作为输入材料。使用 Thermo Fisher Scientific Qubit 4 荧光计和 Quant-iT™ 1X dsDNA 高灵敏度检测试剂盒(Thermo Fisher Scientific,#Q33232)测定从 8 个和 24 个样本的生物学验证运行中获得的文库的 DNA 浓度。平均样品产量为 344.3 ng(+/- 51.5 ng)

为什么这么难以理解我们不...
在尝试理解系统性红斑狼疮 (SLE) 时,我们发现自己处于疾病分类学、致病性导向科学、哲学、经验主义和合格猜想的智力交叉点。科学理论的一个重要结果是,未经批判性研究的科学假设有可能被转变为科学教条 1。这种说法对本研究有影响。我们讨论了两个主要的问题方面。首先,我们必须考虑分类标准的新选择原则 — — 这意味着整合因果关系原则。其次,如果我们想要了解 SLE,就必须实施核心历史数据。这些数据包含与遗传机制相关的独特的、动态变化的 DNA 结构的著名描述。自几十年前发现以来,这些独特的结构在 SLE 研究中大多被忽视。同样,尚无定论的教条数据表明,不同的肾小球配体可被肾炎抗 dsDNA 抗体识别——暴露的染色质片段或固有的膜配体。这些不一致的模型尚未得到比较和系统的研究。本文将深入讨论三个研究领域:(i) SLE 分类标准的选择和作用,该过程必须暗示因果关系原则;(ii) 抗 dsDNA 结构特异性抗体的定义和影响;(iii) 解释狼疮性肾炎的不一致的致病模型。一个精确且至关重要的问题是,SLE 本身是对引发一系列下游效应(标准)的主导统一原因的反应,还是 SLE 代表对多个因果事件随机相互作用的综合反应。这些原则上不同的解释如今并未被正式排除或接受。目前,SLE 可能被视为一种具有表型多样性的疾病,其表现形式独立分离,病因不明,并非单一 SLE 表型所独有。本讨论的重点基本上是

遗传学
Cas,CRISPR 相关;CRISPR,成簇的规律间隔的短回文重复序列;CRISPRa,CRISPR 介导的转录激活;CRISPRi,CRISPR 介导的转录抑制;crRNA,CRISPR RNA;crRNP,CRISPR 核糖核蛋白;dCas9,核酸酶失活 Cas9;DSB,双链断裂;dsDNA,双链 DNA;dsODN,双链寡脱氧核苷酸;gRNA,向导 RNA;H3K27ac,组蛋白 H3 赖氨酸 27 乙酰化;H3K4me1,组蛋白 H3 赖氨酸 4 单甲基化;LAM-PCR,线性扩增介导的 PCR;LSD1,赖氨酸特异性组蛋白去甲基化酶 1;MCP,MS2 外壳蛋白;MOI,感染复数; p65AD,核因子-κB反式激活亚基激活结构域;PAM,原型间隔区相邻基序;RNAi,RNA干扰;scFV,单链可变片段;sfGFP,超折叠GFP;sgRNA,单向导RNA;ssRNA,单链RNA。

新英格兰Biolabs分析证书
在许多DIY基因合成工作流中,用户通过购买合成的dsDNA(例如Gblocks)或准备重叠的寡核体的扩增子来获得用于组装的片段。通常,这些部分具有直接由用于生成碎片的寡核苷酸引起的残余误差。身份酸酶将减少/去除来自扩增子的不匹配/indel(插入/缺失)区域,这些区域源自寡核苷酸化学合成过程中掺入的误差。以下方案增加了经酶校正的DNA池中正确片段的群体,然后允许DNA聚合酶扩增以更准确性和效率富集扩增子。校正和富集步骤的组合增强了组装基因合成的质量,从而用所需的正确的DNA序列产生了较高数量的转化细菌菌落。

HAMILTON NGS STARlet 上的 Oxford Nanopore 连接测序试剂盒 XL V14 自动化为 Nanopore Sequen 生成高质量 DNA 文库
资格设置和结果 为了在 NGS STARlet 上对 Oxford Nanopore SQK-LSK114-XL V14 V1.0 方法进行生物学验证,对 8 个(4 个阳性样本 + 4 个阴性对照)或 24 个样本(22 个阳性样本 + 2 个阴性对照)进行了生物学运行。作为输入材料,1 μg 全长(48 kB)噬菌体 Lambda DNA 用于 8 个样本的运行。对于 24 个样本的运行,1 μg 剪切(9kB)人类基因组 DNA 作为输入材料。使用 Thermo Fisher Scientific Qubit 4 荧光计和 Quant-iT™ 1X dsDNA 高灵敏度检测试剂盒(Thermo Fisher Scientific,#Q33232)测定从 8 个和 24 个样本的生物学验证运行中获得的文库的 DNA 浓度。平均样品产量为 344.3 ng(+/- 51.5 ng)

Authenticase M0689 手册
在许多 DIY 基因合成工作流程中,用户通过购买合成的 dsDNA(例如 gBlocks)或制备重叠寡核苷酸的扩增子来获得用于组装的片段。通常,这些部分具有直接来自用于生成片段的寡核苷酸的残留错误。Authenticase 将减少/去除扩增子中源自寡核苷酸化学合成过程中插入错误的错配/插入/缺失区域。以下方案增加了酶校正 DNA 池中正确片段的数量,随后允许 DNA 聚合酶扩增以更准确和更高效地富集扩增子。校正和富集步骤的结合提高了组装基因合成的质量,从而产生了更多具有所需正确 DNA 序列的转化细菌菌落。
