XiaoMi-AI文件搜索系统
World File Search SystemdsRNA

功能基因组学为物种形成假说提供了新的检验方法
RNA 干扰 (RNAi) 是一种抗病毒真核细胞途径,它在识别细胞质中的 dsRNA 后,靶向并消化相应的 mRNA 链,从而暂时抑制基因表达 [6]。它是一种分子方法,通过将 RNA 分子注入生物体来中和互补的靶 mRNA 分子来改变基因表达。RNAi 沉默机制存在于许多(但不是全部)真核生物中。在进化生物学应用中使用 RNAi 的主要优势是:1)当敲除导致致死时,可以研究必需基因的功能;2)应用于研究难以在胚胎(卵)阶段处理的物种,这是一些替代方法(包括下面讨论的方法)的先决条件。

生物技术通知文件号000197 CVM文件
沉默机制。BG25马铃薯中修饰的第二种预期效应是降低糖的水平并减少酶促变暗(称为“黑点”)。Simplot引入了含有液泡转化酶基因(VINV)和多酚氧化酶基因(PPO)的倒重复段的DNA序列,它们产生DSRNA以降低VINV和PPO的RNA转录水平。VINV基因编码VINV蛋白,该蛋白参与将蔗糖转化为其成分减少糖,而PPO基因编码PPO蛋白,该PPO蛋白氧化酚类化合物可产生深色色素。第三,Simplot引入了来自卵巢结核的改性乙酰乳酸合酶基因(Stmals),该基因编码了stmals蛋白,该蛋白具有对乙酰乳酸合酶(ALS)的耐受性,可抑制除草剂,并用作可选的标记。
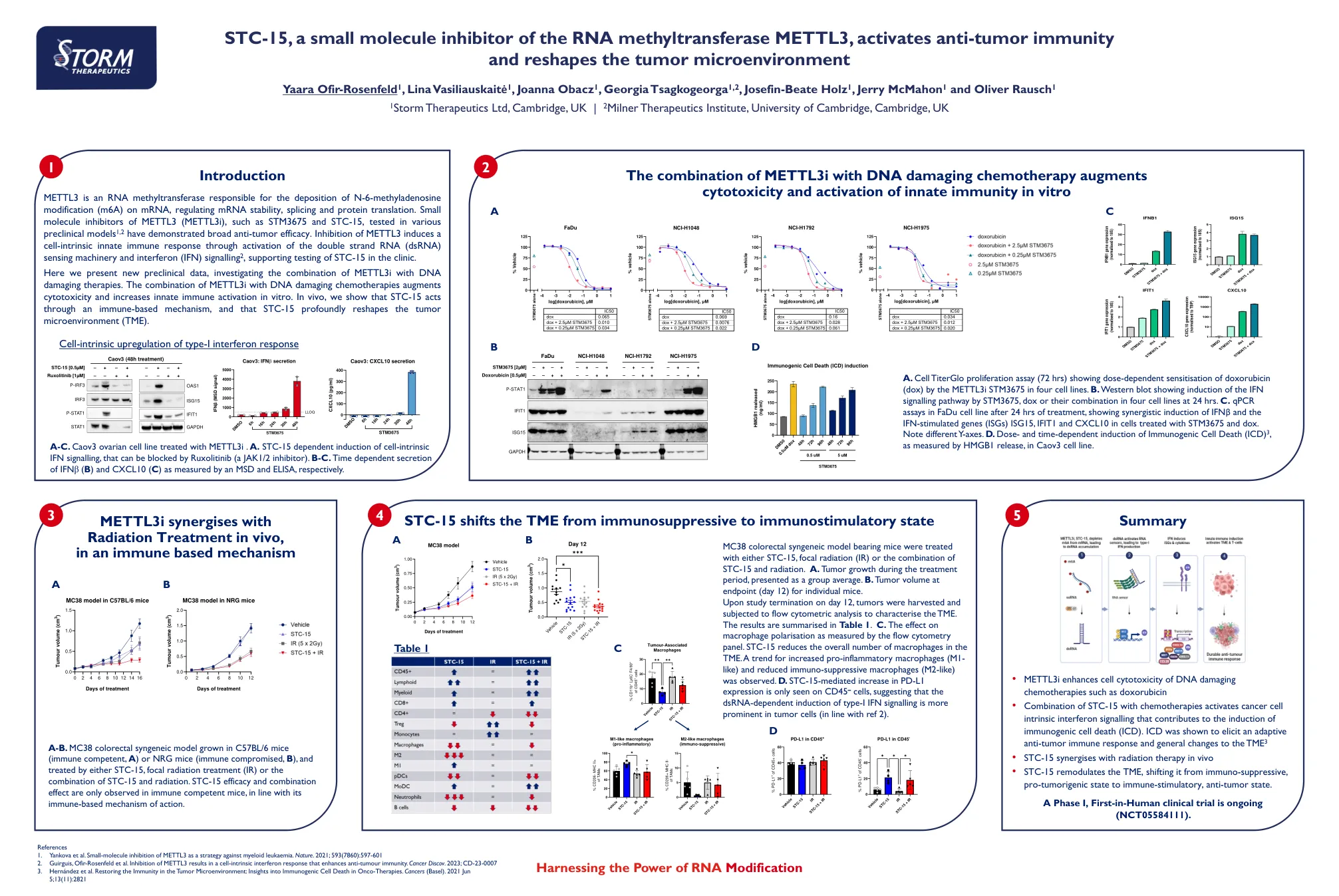
STC-15,一种RNA甲基转移酶METTL3的小分子抑制剂,激活抗肿瘤免疫
mettl3是一种RNA甲基转移酶,负责在mRNA上沉积N-6-甲基二糖苷修饰(M6A),从而调节mRNA稳定性,剪接和蛋白质翻译。在各种临床前模型1,2中测试的METTL3(METTL3I)的小分子抑制剂,例如STM3675和STC-15。抑制METTL3通过激活双链RNA(dsRNA)传感机械和干扰素(IFN)信号2,诱导细胞内用免疫反应,支持诊所中STC-15的测试。在这里,我们提供了新的临床前数据,研究了METTL3I与DNA损害疗法的组合。METTL3I与DNA损害化学疗法的组合增强了细胞毒性并增加了体外先天免疫激活。在体内,我们表明STC-15通过基于免疫的机制起作用,而STC-15深刻重塑了肿瘤微环境(TME)。

一种新颖的定义TLR3激动剂为有效的疫苗佐剂
由Toll样受体3(TLR3)识别的合成双链RNA类似物是疫苗的有吸引力的辅助候选者,尤其是针对细胞内病原体或肿瘤,因为它们具有增强T细胞和抗体反应的能力。尽管Poly(I:C)是具有有效辅助性的代表性DSRNA,但由于异质分子大小,活性不一致,稳定性和毒性不一致,其临床应用受到限制。为了克服这些局限性,我们通过使用PCR偶联的双向体外转录开发了一种新型的基于DSRNA的TSRNA TLR3激动剂(NVT)。琼脂糖凝胶电泳和反相HPLC分析表明,NVT是一个单个275 kDa均匀分子。nvt似乎在6个月的加速存储条件下不受影响,因为其外观,浓度和分子大小不受影响。此外,在良好的实验室实践下对毒性的临床前评估表明,NVT是一种安全物质,没有任何严重毒性迹象。NVT刺激了TLR3,并增加了病毒核酸传感器TLR3,MDA-5和RIG-1的表达。当肌肉内注射到C57BL/6小鼠中时,椭圆蛋白(OVA)和NVT高度增加了树突状细胞(DC)的迁移(DC),巨噬细胞和中性粒细胞与单独使用OVA相比。此外,NVT基本上诱导了直流成熟和激活的表型标记,包括MHC-II,CD40,CD80和CD86以及IFN-B的产生。同时,NVT增加了Th1型T细胞的水平,例如IFN-G + CD4 +和IFN-G + CD8 +细胞,响应OVA刺激。此外,NVT表现出适当的辅助性,因为它升高了OVA特异性IgG,尤其是IgG2C(Th1-型)的较高水平,但IgG1较低(Th2-Type)。总的来说,我们建议具有适当安全性和有效性的NVT是一种新颖而有希望的辅助疫苗,尤其是那些需要T细胞介导的免疫力的疫苗,例如病毒和癌症疫苗。

喷雾诱导的基因沉默:植物特质改善和疾病控制的创新策略
摘要:即使使用最先进的技术,例如基因编辑,现代植物繁殖仍然是一个耗时且昂贵的过程。因此,迫切需要开发植物特质操纵和植物保护的替代方法。RNA干扰(RNAi)是一种由天然存在的双链RNA(DSRNA)和小RNA(SRNA)介导的保守细胞机制,可以靶向mRNA用于破坏或减少转录的mRNA。在这里,我们回顾了基于RNAi的技术的潜力,称为喷雾诱导的基因沉默(SIGS),是在植物或病原体控制中操纵内源基因表达的繁殖的替代或辅助。SIGs可能在减少害虫或病原体影响的情况下特别有用,从而改善生物胁迫并提高作物的农艺性能。关键字:RNA干扰,小RNA,SIGS,DSRNAS
在癌变中,RNA编辑在癌组织中的影响
抽象的RNA编辑是在正常生理过程中观察到的最普遍,最丰富的转录后RNA修饰形式之一,在包括癌症在内的疾病中通常异常。RNA编辑会改变mRNA的序列,使其与源DNA序列不同。编辑的mRNA可以产生与相应基因组编码的蛋白质同工型功能不同的编辑蛋白质同工型。哺乳动物中的主要类型RNA编辑是通过在双链RNA(DSRNA)中或前mRNA转录本中的双链RNA(DSRNA)或发夹中酶脱氨酸对腺苷(A-to-I)的酶促脱氨酸的。催化这些过程的酶属于作用于RNA(ADAR)家族的腺苷脱氨酶。 使用体外癌细胞培养模型获得了与人类疾病相关的RNA编辑景观的绝大多数知识。 但是,这种体外模型的局限性是,获得的结果的生理或疾病相关性不一定是显而易见的。 在这篇综述中,我们专注于讨论在人类癌组织中使用手术切除或从患者临床检索的样品中发现的RNA编辑事件。 我们讨论体内肿瘤中发生的RNA编辑事件如何识别与人类癌症生理学相关的病理信号传导机制,这与癌症进展的不同阶段有关,包括起始,促进,生存,生存,增殖,免疫逃生和转移。催化这些过程的酶属于作用于RNA(ADAR)家族的腺苷脱氨酶。使用体外癌细胞培养模型获得了与人类疾病相关的RNA编辑景观的绝大多数知识。但是,这种体外模型的局限性是,获得的结果的生理或疾病相关性不一定是显而易见的。在这篇综述中,我们专注于讨论在人类癌组织中使用手术切除或从患者临床检索的样品中发现的RNA编辑事件。我们讨论体内肿瘤中发生的RNA编辑事件如何识别与人类癌症生理学相关的病理信号传导机制,这与癌症进展的不同阶段有关,包括起始,促进,生存,生存,增殖,免疫逃生和转移。

miRNA 和 siRNA 对口腔生物群落影响的范例
短核苷酸序列(如 miRNA 和 siRNA)在口腔生物群落研究中引起了广泛关注。miRNA 是一小类非编码 RNA,可调节基因表达以有效调控转录后。相反,siRNA 是 21 – 25 个核苷酸的 dsRNA,通过抑制 mRNA 实现同源依赖性基因沉默,在转录后损害基因功能。本综述重点介绍了 miRNA 在口腔生物群落中的应用,包括口腔癌、牙种植体、牙周病、牙龈成纤维细胞、口腔黏膜下纤维化、放射性口腔黏膜炎、牙髓和口腔苔藓样病。此外,我们还讨论了 siRNA 在上述疾病中的应用,以及 miRNA 和 siRNA 对牙科疾病的各种途径和分子效应物的影响。阐明了 miRNA 和 siRNA 治疗后分子效应物的上调和下调及其对临床环境的影响。因此,上述有关 miRNA 和 siRNA 应用的细节将为学者们提供一个新途径,不仅可以缓解牙科领域的长期问题,还可以开发新的诊断方法。

牡蛎疱疹病毒 1 感染期间的 ADAR 编辑:事实与局限性
摘要 在感染巨牡蛎 (Crassostrea gigas) 的过程中,牡蛎疱疹病毒 1 (OsHV-1) RNA 会通过 A 到 I 的转化进行酶促修饰。与 OsHV-1 RNA 平行的 ADAR1 表达和超编辑活性的增加表明 dsRNA 编辑与抗病毒反应之间存在功能性联系。我们分析了 87 个 RNA 测序数据集,这些数据集来自暴露于 OsHV-1 的免疫致敏、抗性和易感牡蛎,以比较宿主和病毒转录本上的 ADAR 超编辑水平并追踪牡蛎基因上的超编辑。尽管在感染后期病毒 RNA 的编辑有所增加,但宿主 RNA 比病毒 RNA 更容易发生超编辑。一组占牡蛎转录组 0.5% 的基因(包括几个含三部分基序的序列)不断被超编辑。相反,我们鉴定出参与抗病毒反应、miRNA 成熟和表观遗传调控的基因,这些基因仅在特定条件下被过度编辑。尽管技术和生物学瓶颈阻碍了对双壳类“RNA 编辑组”的理解,但现有的工具和技术可以适用于双壳类软体动物。

牡蛎疱疹病毒 1 感染期间的 ADAR 编辑:事实与局限性
摘要 在感染巨牡蛎 (Crassostrea gigas) 的过程中,牡蛎疱疹病毒 1 (OsHV-1) RNA 会通过 A 到 I 的转化进行酶促修饰。与 OsHV-1 RNA 平行的 ADAR1 表达和超编辑活性的增加表明 dsRNA 编辑与抗病毒反应之间存在功能性联系。我们分析了 87 个 RNA 测序数据集,这些数据集来自暴露于 OsHV-1 的免疫致敏、抗性和易感牡蛎,以比较宿主和病毒转录本上的 ADAR 超编辑水平并追踪牡蛎基因上的超编辑。尽管在感染后期病毒 RNA 的编辑有所增加,但宿主 RNA 比病毒 RNA 更容易发生超编辑。一组占牡蛎转录组 0.5% 的基因(包括几个含三部分基序的序列)不断被超编辑。相反,我们鉴定出参与抗病毒反应、miRNA 成熟和表观遗传调控的基因,这些基因仅在特定条件下被过度编辑。尽管技术和生物学瓶颈阻碍了对双壳类“RNA 编辑组”的理解,但现有的工具和技术可以适用于双壳类软体动物。

通过长的非编码RNA(LNCRNA)的转录后调节
从真核基因组中发现了数千种非编码RNA(NCRNA)已彻底改变了生物学的“中心教条”,并转移了对RNA作为调节分子的作用的关注,而不仅仅是基因组信息的传统介体。非编码RNA是不编码蛋白质的转录本,通常根据其平均大小(<或> 200 nt)分类为短或长。在几乎所有生物体中都发现了非编码RNA。其中,长期的非编码RNA(LNCRNA)在许多生物过程中在发育和疾病中起关键作用。自发现以来,lncRNA领域已经爆炸,而lncrnas的新作用不断出现,这使得他们的研究是研究任何水平的基因表达调节的优先事项。本期特刊涵盖了NCRNA场专家的七篇评论论文和一篇原始研究文章,并说明了LNCRNA在转录后层次调节基因表达的主要机制。此文章集提供了其多功能角色的完整概述,并在基因表达和相关细胞过程的调节中提出了额外的复杂性。lncRNA的长度,低表达和缺乏序列保守性经常代表其识别和表征的主要技术限制。在他们的评论中,Carter等。提供了详尽的指南,在硅和低到高的吞吐量实验方法中,以帮助研究人员面对这一挑战。Sadeq等。在扩散的大B细胞淋巴瘤细胞中发现他们还提供了关键的见解,以促进我们对LNCRNA如何参与肿瘤发生的理解[1]。已显示出多种RNA结合蛋白(RBP)与LNCRNA合作以调节基因表达。在评论中,Briata和Gherzi引起了人们对LNCRNA – RBP关联的复杂性的关注[2]。它们说明了LNCRNA-RBP复合物可以控制细胞中所有转录过程的各种机制。讨论了内源性LNCRNA相关的DSRNA结构的耐受性,而病毒衍生的DSRNA触发了一个复杂的防御网络;并进一步研究了自身免疫性疾病和癌症治疗的潜在影响[3]。在他们的综述中,Pisignano和Ladomery描述了LNCRNA有助于调节替代剪接的多种机制,以及它们的作用如何进一步增强了mRNA刺激变体的表达,从而增加了复杂生物体中蛋白质组织的多样性[4]。在更细胞质的环境中,卡拉卡斯和Ozpolat讨论了LNCRNA如何通过控制正常和肿瘤条件下的翻译因子和信号通路来影响mRNA的翻译[5],而Sebastian-Delacruz等。强调了LNCRNA在调节mRNA稳定性和离职率方面的重要性,这是细胞过程和稳态的正确功能的基础[6]。在这方面,在本期特刊中提出的另一项工作中,Munz等。