XiaoMi-AI文件搜索系统
World File Search Systemlei

2025 年第 126 号法案
它建立了巴西疫苗和高价癌症药物的监管框架,并为癌症疫苗的开发、研究、生产、分销和获取制定标准,重点关注科学创新、统一医疗系统(SUS)的普遍获取和公平性,并制定了促进研究、国内生产和国际合作的指导方针。
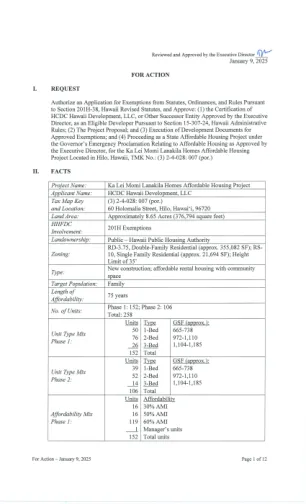
ka lei momi lanakila Homes 201H豁免
B.开发商是一个单一的目的开发实体,由Highridge Costa私有和管理,Highridge Costa是一家专注于经济适用房的房地产开发公司。Highridge Costa专注于为家庭和老年人设计,开发和融资负担得起的公寓。Highridge Costa成立于1994年,曾参与整个美国300个社区的30,000多个经济适用房屋,Highridge Costa在使用联邦第42节LIHTC,退税抵押抵押贷款收入债券,Fannie Mae负担得起的住房和各种州和本地住房计划方面具有经验。Highridge Costa最近参与HHFDC包括690 Pohukaina,Kaahumanu Homes重建,Kokua高级Hale Apartments,Liloa Hale,Pahoa Ridge,Pahoa Ridge,市长Wright Homes Homes Redevelopments,Ka Lei Momi Momi Momi Kapaa和Kapolei Moena contivation(Kapolei Moena)(Hale Moena)(Hale Moena)(Hale Moena)。

2022年12月5日,美国东部时间上午11:07更新了12:19 PM融资单元/Gene TX,由Lei Lei Wu
格林伯格说,Sonothera的平台可以承担他所看到的所有四个,这是当前基因疗法的主要挑战 - 免疫反应,有效负载上限,成本和器官系统的选择。sonothera通过iV与DNA的超声对比剂,然后在目标器官处使用超声探针,在该探针中,声能破坏细胞膜以允许遗传有效载荷进入细胞和细胞核。格林伯格指出,由于裸露的DNA降解很快,因此“在几分钟内”进行了处理和超声检查。

Lei Wang专业知识:我是一个大气动力学家。我的... 生物学未成年人(BIOS)| Purdue EAPS
Lei Wang专业知识:我是一个大气动力学家。 我的研究重点是中纬度大气波的基本动力和变异性,尤其是高影响力的极端天气和气候事件。 我领导着普渡大学的气候和气候动态(WCD)实验室,我还是普渡大学计算跨学科研究生计划(CIGP)的关联教师。 普渡大学地球,大气和行星科学系的现任/最新职位助理教授;哈佛大学博士学位:芝加哥大学,地球物理科学学院,地球和行星科学系博士后研究员,2016年•论文:南方有限振幅波活动的周期性行为Lei Wang专业知识:我是一个大气动力学家。我的研究重点是中纬度大气波的基本动力和变异性,尤其是高影响力的极端天气和气候事件。我领导着普渡大学的气候和气候动态(WCD)实验室,我还是普渡大学计算跨学科研究生计划(CIGP)的关联教师。普渡大学地球,大气和行星科学系的现任/最新职位助理教授;哈佛大学博士学位:芝加哥大学,地球物理科学学院,地球和行星科学系博士后研究员,2016年•论文:南方有限振幅波活动的周期性行为

张磊 2011 年博士论文结题 - 研究探索者
................................................ . ……………………………… ...................................... 123

第355/2024号法案作者:Felipe副手...
自2010年以来,巴西讨论了HPV跟踪测试的实施。与细胞学相比,2010年INCA宫颈癌的“行动计划”的“降低INCA宫颈癌的发病率和死亡率”已经强调了DNA-HPV测试的更高敏感性和较低的特异性,这表明其采用了跟踪,然后在阳性病例中进行细胞学筛查。此外,已经观察到了细胞学检查质量的问题,例如不令人满意的样本的比例,在某些地区,这些样本超过了谁的限制。

卢拉制裁阻止实施新 DPVAT 的法律
经过超过 12 小时的会议,坎皮纳大市议会于昨天凌晨批准了《2025 年年度预算法》(LOA)。该提案确定明年的总预算为 22 亿雷亚尔,并将资源分配到几个优先领域。市卫生基金将获得 6.076 亿雷亚尔,而教育部门的预算为 5.51 亿雷亚尔。公共工程部获得了 2.31 亿雷亚尔的拨款,其次是行政部,获得 9600 万雷亚尔,以及城市服务和环境部,获得 8400 万雷亚尔。市立法部门则将获得 4000 万雷亚尔。剩余资源将根据多年期计划(PPA)和预算指导法(LDO)中定义的规划进行分配。 2025 年 LOA 的一个重要新特点是议员通过强制性修正案。每位议员有权获得市政预算的 0.7%,用于工程、项目或机构支持。这一比例将逐步增加,到 2028 年将达到 1.2%。LOA 的最终文本在财政、预算、金融监督和控制委员会组织的公开听证会上进行了讨论。该委员会由 DEM 议员 Waldeny Santana(照片)组成; Jô Oliveira,来自 PCdoB;来自 PROS 的 Carol Gomes 在此事的评估中发挥了核心作用。在辩论中,议员们强调了民众参与确定预算优先事项的重要性。

Ya-Wen Lei。镀金笼:技术,开发...Ya-Wen Lei。镀金笼:技术,开发...
Lei的重要书籍很好地展示了一个主要国家的理由成为科学和技术而不是人们的福利时会发生什么。 但是,这本书本来可以通过一些历史化来加强。 例如,以儒家伦理和道德的名义,一个继续声称自己是永恒文明瓦斯的承载者的国家如何以科学的名义忘记了人类价值观? 尽管这么小的怪物,但所有学科的中国的学者还是应该阅读这本出色而写得很好的书。 此外,任何思考或从事科学技术在现代社会中的作用的人都将从这项精细的研究中学到很多东西。 其主要贡献是展示国家对控制中国及其社会的意识形态拥抱如何被视为转向全球规范的情况。 取而代之的是,中国需要掌握科学和技术如何独特地变得比实际工具更重要的。他们实施了整个社会的合理化和社会政治控制。 中国只能以自己的方式理解。Lei的重要书籍很好地展示了一个主要国家的理由成为科学和技术而不是人们的福利时会发生什么。但是,这本书本来可以通过一些历史化来加强。例如,以儒家伦理和道德的名义,一个继续声称自己是永恒文明瓦斯的承载者的国家如何以科学的名义忘记了人类价值观?尽管这么小的怪物,但所有学科的中国的学者还是应该阅读这本出色而写得很好的书。此外,任何思考或从事科学技术在现代社会中的作用的人都将从这项精细的研究中学到很多东西。其主要贡献是展示国家对控制中国及其社会的意识形态拥抱如何被视为转向全球规范的情况。取而代之的是,中国需要掌握科学和技术如何独特地变得比实际工具更重要的。他们实施了整个社会的合理化和社会政治控制。中国只能以自己的方式理解。

PL 1118/2024 - 法案 - 1118/2024 - 副议员 Rogerio Morro da Cruz - (121811)
该项目正朝着这个方向发展,将 2 月 20 日设立为地区预防儿童和青少年过早饮酒日。在这一天,将开展教育活动,例如讲座、研讨会和宣传活动,旨在提高人们对儿童和青少年饮酒危害的认识,涉及学校、家庭和整个社区,目的是创造预防文化并鼓励健康行为。

简要回顾 铜基材料作为锂离子电池的阳极和阴极材料 雷刚 1* 和徐春香 2
1 郑州工程学院机电与车辆工程学院,河南郑州 450044,中国 2 郑州工程学院土木工程学院,河南郑州 450044,中国; * 电子邮件:htx510@21cn.com 收稿日期:2020 年 1 月 2 日 / 接受日期:2020 年 2 月 28 日 / 发表日期:2020 年 4 月 10 日 随着对锂离子二次电池能量密度和功率容量的要求越来越高,人们开始寻找容量和性能更好的电极材料。铜基材料因其独特的纳米结构、高电导率和热导率,被认为是改善锂离子电池电化学性能的理想添加剂。综述了铜基纳米材料在电极材料中的应用。本文讨论了铜基纳米复合材料的物理、传输和电化学行为。本文还讨论了铜基纳米复合材料应用面临的挑战及其未来的发展前景。关键词:锂离子电池;铜基材料;纳米复合材料;阳极;阴极 1. 引言