XiaoMi-AI文件搜索系统
World File Search System基因毒性

海报:resDNASEQTM HEK293 和 E1A 片段长度定量分析:基因治疗和疫苗制造中 GMP 批签发的综合解决方案
目的生物制药产品必须限制宿主细胞残留 DNA 污染物,以防止对患者产生基因毒性和免疫毒性风险。现有的残留宿主细胞 DNA 监管指南要求每剂量 ≤10ng,DNA 大小为 200bp 或更低。在病毒载体生产中,衣壳化 DNA 的数量、大小和致癌序列是额外的关注点。为了满足这一需求,赛默飞世尔科技开发了两种旨在满足监管指导的互补检测方法。Applied Biosystems™ resDNASEQ™ 定量 HEK293 DNA 试剂盒可定量总残留宿主细胞 DNA,Applied Biosystems™ resDNASEQ™ E1A DNA 片段长度试剂盒可对针对 E1A 致癌基因的短(86bp)、中(200bp)和长(476bp)片段进行大小分析。

最初发表于:Marion, William;博特彻,斯特芬;鲁伊斯-托雷斯,索尼娅;卢默茨,埃德罗阿尔多;伦丁,凡妮莎;莫里斯,维维安;周,
在这里,我们使用了一种条件性 FA 通路互补系统,其中 FANCA 突变的患者来源的 IPSC 带有可诱导的 FANCA 表达盒。22 我们在造血定向分化系统中使用这些细胞来获得确定的 FA HPC 和同源对照 HPC。23 FANCA 缺陷型、IPSC 衍生的 HPC 表现出与人类 FA 一致的表型,包括对基因毒性应激的敏感性和培养中克隆形成性降低。使用该系统,我们发现 FANCA 缺陷型 HPC 中 p53/p21 轴的激活会阻碍细胞周期进程并驱动终末分化。我们将生长停滞特异性 6 (GAS6) 确定为分化过程中 p53 的靶点,并表明调节 GAS6 信号传导可以挽救 FANCA 缺陷型 HPC 中的造血。该系统克服了使用 IPSC 研究 FA 的挑战,并为进一步研究 FA 病理生物学提供了人类 FA HPC 和同基因对照的可再生来源。

针对功能基因组学的工程化 CRISPR-Cas12a 的高阶组合染色质扰动
多重遗传扰动对于测试编码或非编码遗传元件之间的功能相互作用至关重要。与 DNA 切割相比,使用 CRISPR 干扰 (CRISPRi) 抑制染色质形成可避免基因毒性,并且在混合检测中更有效地扰乱非编码调控元件。然而,目前的 CRISPRi 混合筛选方法通常仅限于每个细胞靶向 1-3 个基因组位点。为了开发一种在功能基因组学筛选中使用 CRISPRi 对基因组位点进行高阶 (> 3) 组合靶向的工具,我们设计了一种 Acidaminococcus Cas12a 变体——称为多重转录干扰 AsCas12a (multiAsCas12a)。 multiAsCas12a 在使用慢病毒转导传递的 CRISPR RNA(crRNA)高阶多路复用阵列进行组合 CRISPRi 靶向时,其表现明显优于最先进的 Cas12a 变体,

果蝇 p53 亚型在生殖系基因组完整性和卵母细胞质量控制方面具有重叠和不同的功能
摘要 人类和其他生物体中的 p53 基因家族成员编码大量蛋白质亚型,其功能大部分尚不明确。以果蝇为模型,我们发现 p53B 亚型主要在生殖细胞中表达,并与 p53A 共定位到亚核体中。然而,只有 p53A 介导生殖细胞和胞体中对电离辐射的凋亡反应。相反,p53A 和 p53B 都是减数分裂 DNA 断裂正常修复所必需的,当减数分裂重组有缺陷时,这种活性更为重要。我们发现在具有持续性 DNA 断裂的卵母细胞中,p53A 也是激活减数分裂粗线期检查点所必需的。我们的研究结果表明,果蝇 p53 亚型具有 DNA 损伤和细胞类型特异性功能,与哺乳动物 p53 家族成员在基因毒性应激反应和卵母细胞质量控制中的作用相似。

使用 GuideScan2 进行全基因组 CRISPR 引导 RNA 设计和特异性分析
基于 CRISPR 的技术已经改变了生命科学,并在治疗学开发中显示出良好的前景 [1],全基因组 CRISPR 筛选通常用于无偏识别各种细胞表型的调节因子。然而,为基于 CRISPR 的基因组扰动设计高效且特异的向导 RNA (gRNA) 带来了计算挑战。不必要的 gRNA 脱靶会导致靶向效率低下以及产生基因毒性,而脱靶信息不完整会导致实验结果的误解 [2]。我们之前开发了 Guide-Scan [3] 用于可扩展的 gRNA 设计,我们和其他人已经证明 GuideScan 在枚举潜在脱靶和估计 gRNA 特异性方面比其他工具更准确 [3, 2]。一个关键的观察结果是,其他 gRNA 设计工具使用的短读比对器虽然对于典型的读取计数量化任务非常有效,但不能详尽地计算次优比对,甚至不能计算多个读取。

CRISPR 染色体碎裂:基因治疗中 CRISPR/Cas9 诱发染色体碎裂的风险
摘要 成簇的规律间隔短回文重复序列 (CRISPR)/Cas9 核酸酶系统已经能够生成疾病模型并开发许多遗传和非遗传疾病的治疗方法。然而,大规模基因组重排的产生引发了人们对 CRISPR/Cas9 核酸酶方法临床应用的安全性担忧。在这些事件中,由于染色体截断而形成的微核和染色体桥可导致局限于一条或几条染色体的大规模基因组重排。这种被称为染色体碎裂的现象最初是在癌细胞中描述的,人们认为它是由有丝分裂过程中染色体分离缺陷或 DNA 双链断裂引起的。在这里,我们将讨论影响 CRISPR/Cas9 诱导的染色体碎裂(以下称为 CRISPR 碎裂)的因素及其结果、表征这些事件的工具以及将其最小化的策略。 关键词:基因组编辑; CRISPR/Cas9;染色体碎裂;基因治疗;基因毒性;微核;染色体不稳定性。

一种正交激活的 CRISPR-Cas13d 纳米前药,可逆转化学耐药性,从而增强化学光动力疗法
通过运输氧化/还原形式的谷胱甘肽及其药物偶联物来改变细胞的氧化还原状态;并且与癌症的不良临床结果(例如预后不良)密切相关。4因此,MRP1 是耐药癌细胞的“致命弱点”之一。5越来越多的证据表明,通过基因沉默方法下调 MRP1 基因可以逆转 MRP1 介导的耐药性。6例如,已发现成簇的规律间隔的短回文重复相关蛋白 9 (CRISPR-Cas9) 技术可以逆转由 ATP 结合盒 (ABC) 转运蛋白介导的 MDR,由于其设计简单、靶区域灵活、编辑效率更高和多路复用,其结果明显高于其他基因编辑技术。 7 – 10 尽管取得了巨大进展,但大多数 CRISPR-Cas9 系统仍然存在一些棘手的问题,包括非靶标基因组改变和基因毒性、Cas9 特异性 T 细胞的潜在免疫风险以及不令人满意的靶向递送。8 为了应对这一挑战,RNA 引导的 VI 型 Cas 蛋白 CRISPR-Cas13d 已被证实可在不改变基因组的情况下敲低靶基因。11,12 重要的是,与 Cas9 蛋白相比,
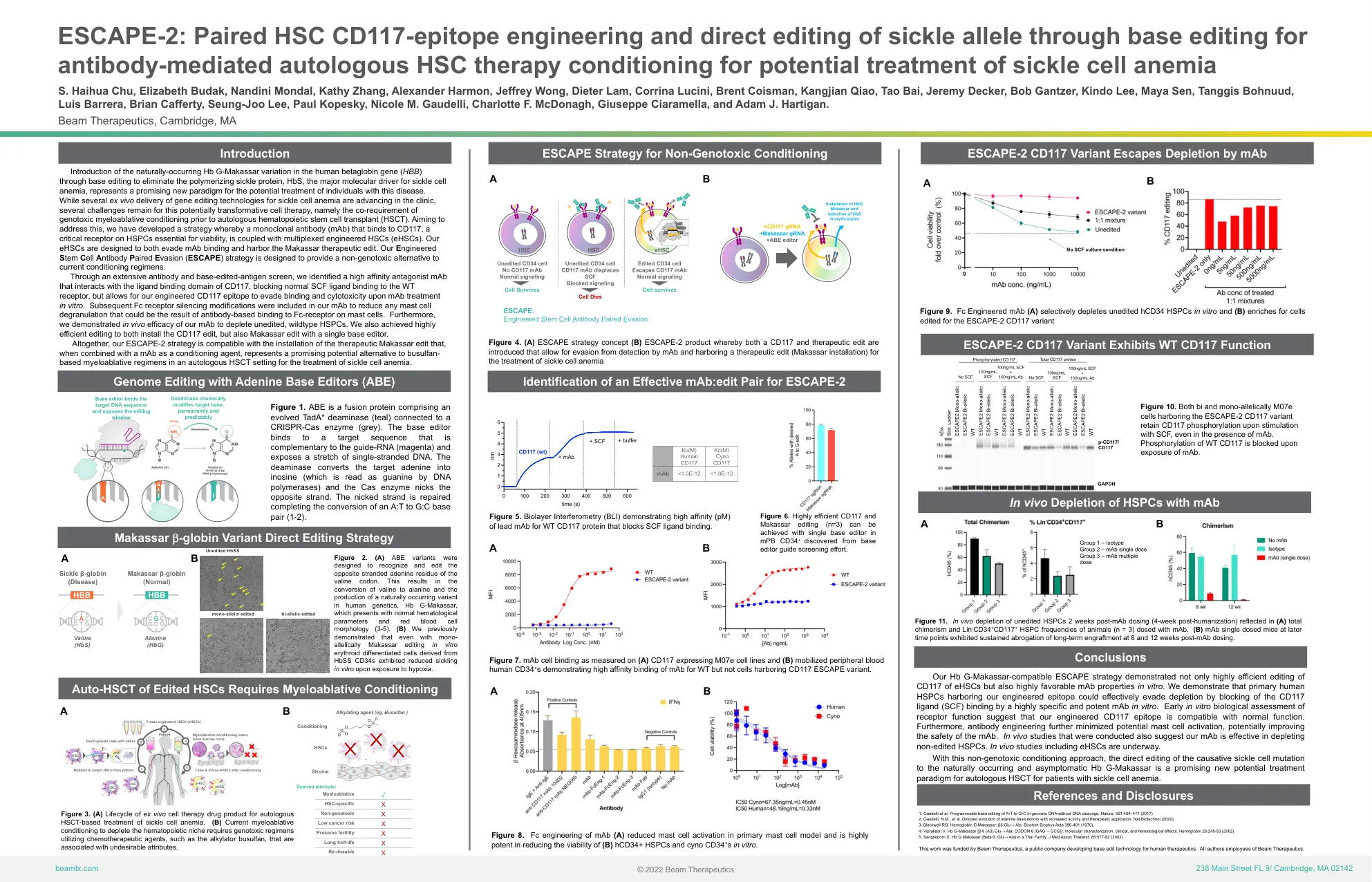
ESCAPE-2:配对 HSC CD117 表位工程和通过碱基编辑直接编辑镰状等位基因,用于抗体介导的自体 HSC 治疗 c
通过碱基编辑在人类β珠蛋白基因 ( HBB ) 中引入天然存在的 Hb G-Makassar 变异,以消除聚合镰状蛋白 HbS(镰状细胞性贫血的主要分子驱动因素),这代表了治疗这种疾病患者的潜在新模式。虽然临床上正在推进几种用于治疗镰状细胞性贫血的体外基因编辑技术,但这种具有潜在变革性的细胞疗法仍然存在一些挑战,即在自体造血干细胞移植 (HSCT) 之前必须进行基因毒性骨髓清除性预处理。为了解决这个问题,我们开发了一种策略,即将一种与 CD117 结合的单克隆抗体 (mAb) 与多重工程化 HSC (eHSC) 结合,CD117 是 HSPC 上对生存至关重要的关键受体。我们的 eHSC 旨在逃避 mAb 结合并携带 Makassar 治疗性编辑。我们的工程干细胞抗体配对逃避(ESCAPE)策略旨在为当前的预处理方案提供一种非基因毒性的替代方案。

化学科学 - RSC 出版
通过运输氧化/还原形式的谷胱甘肽及其药物偶联物来改变细胞的氧化还原状态;并且与癌症的不良临床结果(例如预后不良)密切相关。4因此,MRP1 是耐药癌细胞的“致命弱点”之一。5越来越多的证据表明,通过基因沉默方法下调 MRP1 基因可以逆转 MRP1 介导的耐药性。6例如,已发现成簇的规律间隔的短回文重复相关蛋白 9 (CRISPR-Cas9) 技术可以逆转由 ATP 结合盒 (ABC) 转运蛋白介导的 MDR,由于其设计简单、靶区域灵活、编辑效率更高和多路复用,其结果明显高于其他基因编辑技术。 7 – 10 尽管取得了巨大进展,但大多数 CRISPR-Cas9 系统仍然存在一些棘手的问题,包括非靶标基因组改变和基因毒性、Cas9 特异性 T 细胞的潜在免疫风险以及不令人满意的靶向递送。8 为了应对这一挑战,RNA 引导的 VI 型 Cas 蛋白 CRISPR-Cas13d 已被证实可在不改变基因组的情况下敲低靶基因。11,12 重要的是,与 Cas9 蛋白相比,
利用辐射诱导癌症抗原
摘要:用于治疗癌症的治疗性抗体对晚期患者有效。例如,激活 T 淋巴细胞的抗体可提高许多癌症亚型的存活率。此外,抗体-药物偶联物可有效靶向癌症特异性的细胞毒性药物。本综述讨论了放射诱导抗原,它们是癌症中过度表达的应激调节蛋白。这些可诱导的细胞表面蛋白在细胞对基因毒性应激作出反应时变得易于与抗体结合。主要抗原在所有组织学亚型和几乎所有晚期癌症中均被诱导,但在正常组织中几乎没有表达。通过使用与这些应激调节蛋白特异性结合的治疗性抗体,可利用可诱导抗原。与可诱导抗原 GRP78 和 TIP1 结合的抗体可增强放射治疗在临床前癌症模型中的疗效。细胞毒性药物与抗体的结合进一步改善了癌症反应。本综述重点关注利用放射疗法控制治疗性抗体和抗体-药物偶联物的癌症特异性结合。