XiaoMi-AI文件搜索系统
World File Search System异构化
![具有轴手性和对称性的双苯并[rst]五芬……](/simg/1\122466e03902cc11f82375947dca685fb2d691a5.webp)
具有轴手性和对称性的双苯并[rst]五芬……
探索由两个多环芳烃 (PAH) 单元组成的新型联芳烃是进一步开发具有独特性能的有机材料的重要策略。在本研究中,采用一种高效、通用的方法合成了具有两个苯并[rst]五芬 (BPP) 单元的 5,5′-联苯并[rst]五芬 (BBPP),并通过 X 射线晶体学明确阐明了其结构。BBPP 表现出轴手性,通过手性高效液相色谱法拆分 (M)- 和 (P)-对映体,并通过圆二色光谱法进行研究。根据密度泛函理论计算,这些对映体具有相对较高的异构化能垒,为 43.6 kcal mol − 1。单体 BPP 和二聚体 BBPP 用紫外可见吸收和荧光光谱、循环伏安法和飞秒瞬态吸收光谱进行表征。结果表明,BPP 和 BBPP 均从形式上暗的 S 1 电子态发出荧光,这是通过借用相邻的亮 S 2 态的 Herzberg-Teller 强度实现的。虽然 BPP 表现出相对较低的光致发光量子产率 (PLQY),但由于借用了更大的 S 2 强度,BBPP 表现出显著增强的 PLQY。此外,在不同极性溶剂中进行的光谱研究表明 BBPP 中存在对称性破坏电荷转移。这表明通过适当的分子设计,此类 𝝅 延伸的联芳烃具有很高的单重态裂变潜力。

具有稀有四配位 Ca 中心的丁基锡 Keggin 离子
摘要:烷基锡簇在纳米光刻中用于制造微电子器件。烷基锡 Keggin 家族是整个元素周期表中 Keggin 簇中独一无二的一个,它们似乎更倾向于较低对称性的 β 和 γ 异构体,而不是高度对称的 α 和 ε 异构体。因此,烷基锡 Keggin 家族可能提供有关 Keggin 簇形成和异构化的重要基本信息。我们合成并表征了一种具有四面体 Ca 2+ 中心的新型丁基锡 Keggin 簇,其完整结构为 [(BuSn) 12 (CaO 4 )(OCH 3 ) 12 (O) 4 (OH) 8 ] 2+ (β-CaSn 12 )。合成是一个简单的一步法。广泛的溶液表征包括电喷雾电离质谱、小角度 x 射线散射和多核( 1 H、 13 C 和 119 Sn)NMR,表明 β-CaSn 12 基本上是纯相并且稳定的。这与之前报道的 Na 中心类似物不同,后者总是形成 β 和 γ 异构体的混合物,并且容易相互转化。因此,这项研究澄清了之前对 Na 中心类似物的复杂光谱和晶体学表征的混淆。密度泛函理论计算表明稳定性顺序为 γ-CaSn 12 < γ-NaSn 12 < β-CaSn 12 < β-NaSn 12 ; 类似物总是比 稳定,这与实验一致。这项研究的显著成果包括罕见的四面体 Ca 配位、无 Na 烷基锡簇(对微电子制造很重要)以及对由不同金属阳离子构成的 Keggin 家族的更好理解。

表面张力 - 操纵与可见光的光线 -
摘要:光引起的n = n双键异构化的偶氮元素位于众多应用的核心,从催化,能源储存或药物释放到光遗传学和光电学。While efficient switching between their E and Z states has predominantly relied on direct UV light excitation, a recent study by Klajn and co-workers introduced visible light sensitization of E azoarenes and subsequent isomerization as a tool coined disequilibration by sensitization under confinement (DESC) to obtain high yields of the out-of-equilibrium Z isomer.这种宿主 - 阵线方法仍在高级多组分分子系统中的适用性和功能有限的小型,最小取代的偶氮烯酸含量仍然存在。在此,我们扩展了DESC概念,以引导表面活性剂超分子在空气水接口处。利用可拍摄的芳基唑吡唑两亲物利用我们的专业知识,我们通过可逆的E -Z同源化引起了表面张力和表面过量水的实质性改变。在研究了带电和负电荷的表面活性剂与宿主的结合后,我们发现两种异构体的可见光照射时表面活性差异的程度与直接UV光激发观察到的态度相当。该方法在较大的浓度(从µm到M m)上进行了证明,并且可以使用绿色或红光同样激活,具体取决于选择的敏化剂。在复杂的分子网中,可见光的光电开关敏化的直接实现 - 展示了DESC如何改善现有光响应系统的改善,并允许开发新型应用程序,专门用可见光驱动。

具有稀有四配位 Ca 中心的丁基锡 Keggin 离子
摘要:烷基锡团簇在纳米光刻中用于制造微电子器件。烷基锡 Keggin 家族是整个元素周期表中 Keggin 簇中独一无二的一个;其成员似乎倾向于低对称性的 β 和 γ 异构体,而不是高度对称的 α 和 ε 异构体。因此,烷基锡 Keggin 家族可能为 Keggin 簇的形成和异构化提供重要的基础信息。我们合成并表征了一种具有四面体 Ca 2 + 中心的新型丁基锡 Keggin 簇,其完整结构为 [(BuSn) 1 2 (CaO 4 )- (OCH 3 ) 12 (O) 4 (OH) 8 ] 2+ ( β -CaSn 12 )。该合成是一个简单的一步法。广泛的溶液表征包括电喷雾电离质谱、小角X射线散射和多核( 1 H、 13 C 和 119 Sn)核磁共振,表明β -CaSn 12 基本上是纯相并且稳定的。这与之前报道的Na中心类似物不同,后者总是形成β和γ异构体的混合物,并且容易相互转化。因此,这项研究澄清了之前对Na中心类似物的复杂光谱和晶体学表征的混淆。密度泛函理论计算显示以下稳定性顺序:γ -CaSn 12 < γ -NaSn 12 < β - CaSn 12 < β -NaSn 12。β类似物总是比γ类似物更稳定,这与实验一致。本研究的显著成果包括罕见的四面体 Ca 配位、无 Na 烷基锡簇(对微电子制造很重要)以及对由不同金属阳离子构成的 Keggin 家族的更好理解。■ 简介

2024-2025 研讨会摘要 - 化学
Thanh Do,分析化学 驯服离子赛道上的构象异质性 大约 85% 的人类蛋白质组无法通过传统的小分子进行治疗。潜在的药物必须足够大且足够灵活,才能与大的凹槽状结合位点结合,或结合在两种蛋白质的界面上。环孢菌素是一类 N-甲基化的大环肽,它挑战了基于结构药物设计的传统观点。尽管环孢菌素 A (CycA) 自 1983 年以来彻底改变了器官移植领域,但针对不同靶点设计类似药物的尝试均未成功,这表明在 N-甲基化的作用和构象异质性在环孢菌素化学中的功能方面存在知识差距。环孢菌素由于 N-甲基化而具有灵活性,每次顺式/反式酰胺异构化都会改变分子的构象和物理化学性质。 CycA 可以与多个靶标(目前已知两个)结合,结合状态不同,这表明结合状态取决于靶标。先前的研究表明,结合状态(与已知靶标)在溶液中以次要构象异构体的形式存在。这表明环孢菌素可能通过反向诱导契合模型与其靶标结合,其中配体改变其构象以适应结合位点。因此,环孢菌素可以结合的靶标数量可能与其可以采用的可能构象异构体数量成正比。因此,为了充分了解环孢菌素的生化特性,我的实验室致力于准确探测 CycA 和 CycA 类似物的主要和次要构象异构体,使用多种技术,包括实验(X 射线/中子晶体学、离子迁移质谱、2D-NMR、离子光谱)和计算方法。我们发现了一个由二价离子调节的复杂构象网络和动力学。

利用荧光蛋白检测生物胶中的断裂
在聚合物机械化学领域 [10,11],OFP [12,13] 可以实现光学可视化,并监测不同材料体系(从传统的热固性材料和热塑性材料 [14–18] 到蛋白质)内不同长度尺度上的机械诱导事件。[19–23] 在机械生物学领域也可以找到类似的概念。[24–27] 在施加力时,OFP 会发生构象、构型或组成键异构化反应,从而改变其在吸收、荧光或化学发光方面的光学性质。[28] 材料科学中高分辨率显微镜技术的出现甚至使我们能够追踪亚微米尺度的宏观材料损伤。[29–37] 因此,OFP 有助于开发具有改进性能的材料方法。 [38] 尽管 OFP 已成功用于研究合成和生物大分子材料的损伤,但令人惊讶的是,尚未使用 OFP 研究粘合剂的失效。现有的研究粘合剂疲劳和断裂的方法[39]包括目视检查、[40] X 射线光电子能谱、[41,42] 质谱 (MS)、[43,44] 傅里叶变换红外光谱、[42,45] 和接触角测量。[42] 然而,这些技术都无法对胶水成分的机械状态提供空间分辨的光学反馈。我们在此报道了一种由阳离子力响应蛋白 FRET 对和阴离子芳香族表面活性剂的静电共聚形成的生物胶。[46,47] 因此,我们将 FRET 供体荧光团连接到力响应的 FRET 受体荧光蛋白。在机械测试过程中,施加力会改变 FRET 效率,从而改变发射光谱以及供体荧光寿命。我们使用这些蛋白质粘合剂粘合高能和低能表面,以对其断裂行为进行详细的光学分析。机械损伤

JJ Vittal的出版物 - 新加坡
评论,科学评论和观点17。J.J. Vittal,“ [2+2]光载载反应是一种通过机械化学研磨来监测固态分子运动的工具”,J。PhotoChem。 光二醇。 c:光化学。 Rev。 57(2023)100636 16。 Y.-L。 Li,A.-J。 li,S.-L。 Huang,J。J. Vittal,G.-Y. Yang,“光催化的polypyridyl ru(II)或环数IR(III)官能化结构”,Chem。 Soc。 Rev。 52(14)(2023)4725-4754 15。 G.K. Kole,J.J。 Vittal,“固态和溶液中环丁烷配体的异构化”,J。Ind. 化学。 Soc。 ,99(9)(2022)100630 14。 B.B. Rath,J.J。 Vittal,“表现出[2 + 2]光载体反应和动态效应的光电反应晶体”,Acc。 化学。 res。 55(10)(2022),1445-1455 13。 G. Chakraborty,I.H。 Park,R。Medishetty,J.J。 Vittal,“ T wo二维金属有机框架材料:综合,结构,性质和应用”,化学。 修订版 ,121(7)(2021)3751-3891 12。 M. Gupta,J.J。 Vittal,“控制MOF的互穿和结构转化的控制”,坐标。 化学。 修订版 ,435(2021)213789(邀请评论)11。 J. J. Vittal,H.S。 quah,“工程固态的固态结构转换”坐标。 化学。 修订版 ,342(2017)1-18(邀请评论)10。 9。 和债券。 157(2014)105-144。J.J. Vittal,“ [2+2]光载载反应是一种通过机械化学研磨来监测固态分子运动的工具”,J。PhotoChem。光二醇。c:光化学。Rev。 57(2023)100636 16。 Y.-L。 Li,A.-J。 li,S.-L。 Huang,J。J. Vittal,G.-Y. Yang,“光催化的polypyridyl ru(II)或环数IR(III)官能化结构”,Chem。 Soc。 Rev。 52(14)(2023)4725-4754 15。 G.K. Kole,J.J。 Vittal,“固态和溶液中环丁烷配体的异构化”,J。Ind. 化学。 Soc。 ,99(9)(2022)100630 14。 B.B. Rath,J.J。 Vittal,“表现出[2 + 2]光载体反应和动态效应的光电反应晶体”,Acc。 化学。 res。 55(10)(2022),1445-1455 13。 G. Chakraborty,I.H。 Park,R。Medishetty,J.J。 Vittal,“ T wo二维金属有机框架材料:综合,结构,性质和应用”,化学。 修订版 ,121(7)(2021)3751-3891 12。 M. Gupta,J.J。 Vittal,“控制MOF的互穿和结构转化的控制”,坐标。 化学。 修订版 ,435(2021)213789(邀请评论)11。 J. J. Vittal,H.S。 quah,“工程固态的固态结构转换”坐标。 化学。 修订版 ,342(2017)1-18(邀请评论)10。 9。 和债券。 157(2014)105-144。Rev。57(2023)100636 16。Y.-L。 Li,A.-J。 li,S.-L。 Huang,J。J. Vittal,G.-Y. Yang,“光催化的polypyridyl ru(II)或环数IR(III)官能化结构”,Chem。 Soc。 Rev。 52(14)(2023)4725-4754 15。 G.K. Kole,J.J。 Vittal,“固态和溶液中环丁烷配体的异构化”,J。Ind. 化学。 Soc。 ,99(9)(2022)100630 14。 B.B. Rath,J.J。 Vittal,“表现出[2 + 2]光载体反应和动态效应的光电反应晶体”,Acc。 化学。 res。 55(10)(2022),1445-1455 13。 G. Chakraborty,I.H。 Park,R。Medishetty,J.J。 Vittal,“ T wo二维金属有机框架材料:综合,结构,性质和应用”,化学。 修订版 ,121(7)(2021)3751-3891 12。 M. Gupta,J.J。 Vittal,“控制MOF的互穿和结构转化的控制”,坐标。 化学。 修订版 ,435(2021)213789(邀请评论)11。 J. J. Vittal,H.S。 quah,“工程固态的固态结构转换”坐标。 化学。 修订版 ,342(2017)1-18(邀请评论)10。 9。 和债券。 157(2014)105-144。Y.-L。 Li,A.-J。li,S.-L。 Huang,J。J. Vittal,G.-Y.Yang,“光催化的polypyridyl ru(II)或环数IR(III)官能化结构”,Chem。Soc。Rev。 52(14)(2023)4725-4754 15。 G.K. Kole,J.J。 Vittal,“固态和溶液中环丁烷配体的异构化”,J。Ind. 化学。 Soc。 ,99(9)(2022)100630 14。 B.B. Rath,J.J。 Vittal,“表现出[2 + 2]光载体反应和动态效应的光电反应晶体”,Acc。 化学。 res。 55(10)(2022),1445-1455 13。 G. Chakraborty,I.H。 Park,R。Medishetty,J.J。 Vittal,“ T wo二维金属有机框架材料:综合,结构,性质和应用”,化学。 修订版 ,121(7)(2021)3751-3891 12。 M. Gupta,J.J。 Vittal,“控制MOF的互穿和结构转化的控制”,坐标。 化学。 修订版 ,435(2021)213789(邀请评论)11。 J. J. Vittal,H.S。 quah,“工程固态的固态结构转换”坐标。 化学。 修订版 ,342(2017)1-18(邀请评论)10。 9。 和债券。 157(2014)105-144。Rev。52(14)(2023)4725-4754 15。G.K. Kole,J.J。 Vittal,“固态和溶液中环丁烷配体的异构化”,J。Ind.化学。Soc。,99(9)(2022)100630 14。B.B.Rath,J.J。 Vittal,“表现出[2 + 2]光载体反应和动态效应的光电反应晶体”,Acc。 化学。 res。 55(10)(2022),1445-1455 13。 G. Chakraborty,I.H。 Park,R。Medishetty,J.J。 Vittal,“ T wo二维金属有机框架材料:综合,结构,性质和应用”,化学。 修订版 ,121(7)(2021)3751-3891 12。 M. Gupta,J.J。 Vittal,“控制MOF的互穿和结构转化的控制”,坐标。 化学。 修订版 ,435(2021)213789(邀请评论)11。 J. J. Vittal,H.S。 quah,“工程固态的固态结构转换”坐标。 化学。 修订版 ,342(2017)1-18(邀请评论)10。 9。 和债券。 157(2014)105-144。Rath,J.J。 Vittal,“表现出[2 + 2]光载体反应和动态效应的光电反应晶体”,Acc。化学。res。55(10)(2022),1445-1455 13。G. Chakraborty,I.H。 Park,R。Medishetty,J.J。 Vittal,“ T wo二维金属有机框架材料:综合,结构,性质和应用”,化学。 修订版 ,121(7)(2021)3751-3891 12。 M. Gupta,J.J。 Vittal,“控制MOF的互穿和结构转化的控制”,坐标。 化学。 修订版 ,435(2021)213789(邀请评论)11。 J. J. Vittal,H.S。 quah,“工程固态的固态结构转换”坐标。 化学。 修订版 ,342(2017)1-18(邀请评论)10。 9。 和债券。 157(2014)105-144。G. Chakraborty,I.H。Park,R。Medishetty,J.J。 Vittal,“ T wo二维金属有机框架材料:综合,结构,性质和应用”,化学。 修订版 ,121(7)(2021)3751-3891 12。 M. Gupta,J.J。 Vittal,“控制MOF的互穿和结构转化的控制”,坐标。 化学。 修订版 ,435(2021)213789(邀请评论)11。 J. J. Vittal,H.S。 quah,“工程固态的固态结构转换”坐标。 化学。 修订版 ,342(2017)1-18(邀请评论)10。 9。 和债券。 157(2014)105-144。Park,R。Medishetty,J.J。 Vittal,“ T wo二维金属有机框架材料:综合,结构,性质和应用”,化学。修订版,121(7)(2021)3751-3891 12。M. Gupta,J.J。 Vittal,“控制MOF的互穿和结构转化的控制”,坐标。 化学。 修订版 ,435(2021)213789(邀请评论)11。 J. J. Vittal,H.S。 quah,“工程固态的固态结构转换”坐标。 化学。 修订版 ,342(2017)1-18(邀请评论)10。 9。 和债券。 157(2014)105-144。M. Gupta,J.J。 Vittal,“控制MOF的互穿和结构转化的控制”,坐标。化学。修订版,435(2021)213789(邀请评论)11。J. J. Vittal,H.S。 quah,“工程固态的固态结构转换”坐标。 化学。 修订版 ,342(2017)1-18(邀请评论)10。 9。 和债券。 157(2014)105-144。J. J. Vittal,H.S。quah,“工程固态的固态结构转换”坐标。化学。修订版,342(2017)1-18(邀请评论)10。9。和债券。157(2014)105-144。M. J. Werny,J.J。 Vittal,“调节三个多晶型物中的热和体性行为”,IUCRJ,4(2017)202-203(受邀的科学评论)。 R. Medishetty,J.J。 Vittal,“光化学反应的金属有机框架”,用于光子学应用的金属有机框架(编辑:B。Chen和G. Qian),结构。 (被邀请)8。 G.K. Kole,J.J。 Vittal,“固态反应性,涉及协调聚合物的结构转化”,化学。 Soc。 修订版 42(4)(2013)1755-1775,(邀请了Werner Issue的评论)M. J. Werny,J.J。 Vittal,“调节三个多晶型物中的热和体性行为”,IUCRJ,4(2017)202-203(受邀的科学评论)。R. Medishetty,J.J。 Vittal,“光化学反应的金属有机框架”,用于光子学应用的金属有机框架(编辑:B。Chen和G. Qian),结构。 (被邀请)8。 G.K. Kole,J.J。 Vittal,“固态反应性,涉及协调聚合物的结构转化”,化学。 Soc。 修订版 42(4)(2013)1755-1775,(邀请了Werner Issue的评论)R. Medishetty,J.J。 Vittal,“光化学反应的金属有机框架”,用于光子学应用的金属有机框架(编辑:B。Chen和G. Qian),结构。(被邀请)8。G.K. Kole,J.J。 Vittal,“固态反应性,涉及协调聚合物的结构转化”,化学。Soc。修订版42(4)(2013)1755-1775,(邀请了Werner Issue的评论)
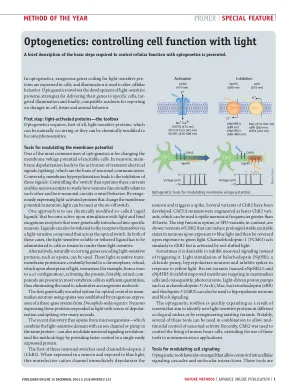
光遗传学:用光控制细胞功能
调节膜电位的工具 光遗传学最常见的用途之一是改变可兴奋细胞的膜电位。在神经元中,膜去极化会导致瞬态电信号(脉冲)的激活,这是神经元通讯的基础。相反,膜超极化会导致这些信号的抑制。控制操作这些电流的“开关”使神经科学家能够研究神经元在功能上如何相互关联以及神经元回路如何控制行为。通过外源表达改变神经元膜电位的光激活蛋白,光可以用作开关。一种方法是使用化学修饰的所谓“笼状配体”,这些配体在光刺激下变得活跃并与通过基因引入特定神经元的外源性受体结合。配体也可以通过充当光开关的光敏化合物与受体本身相连。在这两种情况下,都必须将光敏的可溶性或束缚配体注入细胞或组织,使它们对光敏感。或者,可以使用编码光敏蛋白(如视蛋白)的天然基因。这些光敏跨膜蛋白与发色团视网膜共价结合,视网膜吸收光后发生异构化(例如,从反式变为顺式构型),从而激活蛋白质。值得注意的是,视网膜化合物在大多数脊椎动物细胞中含量充足,因此无需注入外源分子。第一个利用视蛋白进行哺乳动物神经元光学控制的遗传编码系统是通过外源表达果蝇的三基因系统建立的。表达这些蛋白质的神经元对光的反应是数秒内的去极化和尖峰波。最近发现,微生物中的视蛋白(将光敏域与同一蛋白质中的离子通道或泵相结合)也可以调节神经元信号,通过在单个易于表达的蛋白质中提供更快的控制,彻底改变了该方法。这些神经元开关中的第一个使用了通道视紫红质-2 (ChR2)。当在神经元中表达并暴露于蓝光时,这种非选择性阳离子通道会立即使神经元去极化

补充信息,Barends 等人。
补充参考文献 1. Lincoln, CN, Fitzpatrick, AE 和 van Thor, JJ 光活性黄色蛋白飞秒激发下的光异构化量子产率和非线性截面。Phys. Chem. Chem. Phys. 14 , 15752-15764 (2012)。 2. Kim, JE, Tauber, MJ 和 Mathies, RA 视觉中波长依赖性的顺反异构化。Biochemistry 40 , 13774-13778 (2001)。 3. Shoeman, RL, Hartmann, E. 和 Schlichting, I. 生长和制造纳米和微晶体 Nat Protoc 正在印刷中 (2022)。 4. Groot, ML, vanGrondelle, R., Leegwater, JA 和 vanMourik, F. 绿色植物和细菌红细菌光系统 II 反应中心的自由基对量子产率。亚皮秒脉冲下的饱和行为。J. Phys. Chem. B 101 , 7869-7873 (1997)。5. Claesson, E. 等人。飞秒 X 射线激光捕获的光敏色素蛋白的一级结构光响应。eLife 9 , e53514 (2020)。6. Sugahara, M. 等人。油脂基质作为用于序列晶体学的多功能蛋白质载体。自然方法 12 , 61-3 (2015)。7. Li, H. 等人。使用时间分辨的串行飞秒晶体学捕捉光系统 II 从 S1 到 S2 转变的结构变化。IUCrJ 8,431-443 (2021)。8. Grünbein, ML 等人。通过串行飞秒晶体学进行超快泵浦探测实验的照明指南。自然方法 17,681-684 (2020)。9. Nogly, P. 等人。飞秒 X 射线激光捕获细菌视紫红质中的视网膜异构化。科学 361,eaat0094 (2018)。10. Falahati, K.、Tamura, H.、Burghardt, I. 和 Huix-Rotllant, M. 通过非绝热量子动力学实现肌红蛋白中的超快一氧化碳光解和血红素自旋交叉。 Nat Commun 9 , 4502 (2018)。11. Barends, TR 等人。直接观察配体解离后 CO 肌红蛋白中的超快集体运动。Science 350 , 445-50 (2015)。

Hawkmoth信息素转导涉及G蛋白 - ...
抽象的进化压力适应了昆虫化学效应,以适应其各自的生理需求和生态壁ni的任务。孤独的夜间飞蛾依靠他们的急性嗅觉在晚上找到伴侣。通过大多数未知的机制,以最大的灵敏度和高时间分辨率检测到信息素。虽然昆虫嗅觉受体的逆拓扑和与嗅觉受体共感染者的异构化表明通过气味门控受体 - 离子通道复合物的离子型转导,但矛盾的数据提出了扩增的G-protein-G-protein - 耦合的转导。在这里,我们在特定时间中使用了男性甘达·塞克斯塔·霍克莫斯(Manduca Sexta Hawkmoths)的信息素敏感性的体内尖端录制(REST与活动与活动)。由于嗅觉受体神经元在其信息素响应的三个连续时间窗口中区分了信号参数(phasic; tonic; tonic;晚期,持久),因此分别分析了各自的响应参数。G蛋白的破坏 - 偶联的转导和磷脂酶C的阻滞减少并减慢了霍克莫斯活动阶段的阶段反应成分,而不会影响活动和休息期间的任何其他响应。使用细菌毒素阻止Gαo或持续激活GαS的Gα亚基的使用细菌毒素的持续激活影响了变质的信息素反应,而靶向GαQ和Gα12/13的毒素却无效。 因此,可以通过考虑昼夜节律时间和独特的气味响应成分来解决有关昆虫嗅觉的差异。使用细菌毒素的持续激活影响了变质的信息素反应,而靶向GαQ和Gα12/13的毒素却无效。因此,可以通过考虑昼夜节律时间和独特的气味响应成分来解决有关昆虫嗅觉的差异。与这些数据一致,磷脂酶Cβ4的表达取决于Zeitgeber时间,这表明昼夜节律调节的代谢素信息素转导级联级联反应最大化霍克莫斯活性阶段的信息素转导的敏感性和时间分辨率。