XiaoMi-AI文件搜索系统
World File Search System异构化
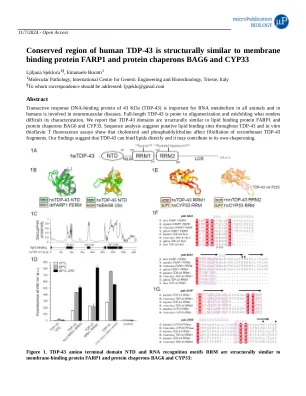
人类TDP-43的保守区域在结构上类似于膜结合蛋白FARP1和蛋白质伴侣Bag6和Cyp33
1a人类TDP-43(HSTDP-43)的示意图:NTD-氨基末端结构域,NLS-核定位信号,RRM-RNA识别基序,LCR-low复杂性区域;在RRM1中类似PIASE的序列和假定的聚集和RRM2中的纤维化启动序列被证明,并以粉红色显示顺式P225。1B HSTDP-43 NTD结构域与斑马鱼Farp1的Ferm结构域以及Dali产生的人类Bag6的泛素样域。1C的HSTDP-43残基的溶解倾向为绘制的TDP-43序列绘制的脂质结合区域无序;预测的脂质结合区域无序表示为黑色矩形,并根据HSTDP-43氨基酸序列编号。重组的1d噻铁黄素T荧光在37°C或65°C下在胆固醇(C)和磷脂酰胆碱(PC)的情况下在生理温度下或在65°C下在生理温度下或65°C下在生理温度下或65°C下在生理温度下孵育的HSTDP-43构建体和对照样品;误差线表示来自一式三份实验的平均值的标准误差。1E HSTDP-43 RRM1和小鼠TDP-43 RRM2主题达利生成的叠加到HSCYP33 RRM域的3D模型; MMTDP43 RRM2顺式Proline P225标有粉红色的星号。1f欧米茄生成的人,小鼠,鸡肉和鱼Farp1和TDP-43的多个序列比对,以及Zebra Fish Ferm域的二级结构元素相对于多个序列对齐信息的二级结构元素的二级结构元素;白色和黑色钻石分别代表了TDP-43和FARP1中的假定或实验确认的脂质结合残基。1G人和小鼠CYP33 RRM和PPIASE结构域的多个序列对齐,以及人和小鼠TDP-43 RRM1和RRM2基序; HSTDP-43 RRM1或HSCYP33 PPIASE域的二级结构元素的ESPRIPT生成的渲染相对于多个序列比对信息; TDP-43 rrm2中的顺式脯氨酸用粉红色的星号表示,并且在所有排列序列中,粉红色矩形突出显示了该位置。 CYP33参与底物结合的残基用白色球表示,其中一些与肽基prolyl prolyl cis-Trans异构化的HSCYP33残基由黑色球体表示,而催化HSCYP33 S239不包括由于空间限制而包括。

腔量子电动力学可以启用para
摘要:将分子耦合到光腔内的量化辐射场已显示出巨大的前景,可以改变化学反应性。在这项工作中,我们表明,可以通过将反应与腔反应强,产生正骨 - 或para取代的产物而不是元产品来从根本上改变硝基苯的基础选择性。重要的是,这些是从腔体以外的同一反应中获得的产物。最近开发的AB从头算法用于理论上计算阳离子卫星中间体的相对能量,这表明所有产品的动力学优选的溴化位点。对腔内和外部的蜂巢中间体的地下电子密度进行分析,我们演示了强耦合如何引起分子电荷分布的重组,这又导致不同的溴化位点直接取决于空腔条件。总体而言,此处介绍的结果可用于了解腔体从机械的角度使用对基态化学反应性的变化,并将前沿理论模拟与最先进但现实的实验腔条件直接连接。■将耦合分子偶联到光腔内的量化辐射场中产生一组光子 - 物质杂种态,称为polaritons。这些极化状态通过调整物质的特性以及光子的特性来以一般和便捷的方式改变化学反应性。23请注意,尽管将极化子用于新的化学的理论预测广泛地,但1已在实验上证明的很大程度上与北极星修饰的反应动力学有关。例如,富尔吉德或类似分子的电子激发态之间的集体耦合以及光腔内量化的光子模式,所谓的电子强耦合(ESC),以增强或抑制光化异构化反应。2,3在另一个示例中,振动激发共同与微腔的光子激发(通常称为振动强耦合(VSC))共同耦合,导致化学动力学可以增强4、5或抑制。6-8在这两个集体耦合方案中,反应的动力学发生了变化,但重要的是,与腔体以外的相同反应相比,没有生成新的产品。最近的理论研究1,9表明,可以通过将分子的电子状态与空腔光子模式耦合来显着修改分子系统的基态。10-20,特别是,已经表明,腔体可以修改Diels- alder反应的内部/EXO选择性,21,22修改了地面质子转移反应屏障和驱动力15,16,并选择性地控制点击反应的乘积。

激发态的量子保真度磁化率......
弯曲振动自由度的研究得益于其二维特性和两个明确的物理极限——线性和弯曲配置——以及中间配置——准线性物种,其特点是大振幅运动,使其具有丰富的光谱特征[1]。正或非单调的非谐性,后者与非刚性分子的 Birge-Sponer 图中 Dixon 凹陷的出现有关[2],以及由于跨越线性壁垒附近的状态波函数中线性和弯曲特征的混合而导致的异常旋转光谱[3,4],是准线性物种光谱中最显著的光谱特征。光谱方法的重大进步和发展使得人们能够通过实验获得多种分子物种的高弯曲泛音。通过这种方式,我们有可能获得实验光谱信息,从而研究能量接近线性势垒的系统 [5,6]。水 [7] 和 NCNCS [8–10] 的研究结果具有特别重要的意义。近年来,量子单值化概念最初由 Cushman 和 Duistermaat [11] 提出,后由 Child [12] 重新研究,对系统中的状态分配有很大帮助。由于状态与线性势垒的接近性,波函数的复杂性妨碍了正确的状态标记 [5–8,13]。这是从经典力学中借用的概念,它依赖于拓扑奇点,当系统能量大到足以探测局部鞍点或最大值时,就会发生拓扑奇点,从而阻止定义全局作用角变量 [14]。非刚性分子弯曲振动的理论建模需要特殊的工具,因为大振幅振动自由度会强烈耦合振动和转动自由度。Hougen-Bunker-Johns 弯曲哈密顿量 [15] 是该领域的一项开创性工作。这项工作后来扩展到半刚性弯曲哈密顿量 [16] 和一般半刚性弯曲哈密顿量 [17]。基于上述发展而产生的 MORBID 模型 [18] 目前是分析非刚性分子光谱的标准方法,其中需要同时考虑转动和振动自由度,以便建模实验项值并分配量子标签。代数方法,尤其是振动子模型,是分子光谱建模的传统积分微分方法的替代方法。该模型基于对称性考虑,并严重依赖于李代数的性质[ 19 ]。振子模型 (VM) 属于一类模型,该类模型将 U(n+1) 代数指定为 n 维问题的动力学或谱生成代数 [20]。类似的模型已成功应用于强子结构 [21,22] 和原子核 [23–25] 的建模。在 Iachello 引入的原始振子模型形式中,双原子分子种类的回旋振动激发被视为集体玻色子激发 [26],由于相关自由度的矢量性质,动力学代数为 U(3+1)=U(4) [25,27]。弯曲振动的二维性质以及简化振子模型形式以有效处理多原子系统的需要,自然而然地导致了二维极限振子模型(2DVM)的制定[28,29]。2DVM 定义的形式能够模拟弯曲自由度的线性和弯曲极限情况,以及表征中间情况的大振幅模式[30-33]。本研究中使用的代数哈密顿量的四体算符的扩展已于最近发表[34]。2DVM 还用于耦合弯曲器[28,35-37]、拉伸弯曲相互作用[38-41]和异构化反应中的过渡态[42]的建模。