XiaoMi-AI文件搜索系统
World File Search System性二态




基于量子纠缠的远程态制备研究进展
[42] Ra Y S,Dufour A,Walschaers M等。多模光场的非高斯量子状态[J]。自然物理学,2020,16(2):144-147。[43] Asavanant W,Yu S,Yokoyama S等。生成时间 - 域 - 多路复用两个维群集状态[J]。Science,2019,366(6463):373-376。[44] Larsen M,Guo X,Breum C等。确定性生成两个维簇状态[J]。Science,2019,366(6463):369-372。[45] Aasi J,Abadie J,Abbott B P等。使用挤压的光态[J]增强了LIGO重力波检测器的灵敏度。自然光子学,2013,7(8):613-619。[46] Yonezawa H,Furusawa A.连续 - 可变的量子信息处理,挤压光态[J]。光学和光谱学,2010,108(2):288-296。[47] Takeda S,Furusawa A.朝向大 - 比例断层 - 耐受性光子量子计算[J]。APL Photonics,2019,4(6):060902。[48]秦忠忠,王美红,马荣,等。压缩态光场及其应用研究[J]。激光与光电子学进展,2022,59(11):1100001。QIN Z Z,Wang M H,Ma R等。挤压光及其应用的进展[J]。激光和光电进度,2022,59(11):1100001。[49] Mari A,Eisert J.阳性Wigner函数呈现量子计算有效的经典模拟[J]。物理评论来信,2012,109(23):230503。[50] Xiang Y,Kogias I,Adesso G等。物理评论A,2017,95(1):010101。多部分高斯转向:一夫一妻制约束和量子加密应用[J]。[51] Xiang Y,Liu S H,Guo J J等。分销和

性二态社会行为的神经控制
在社会行为的神经控制中的快速进步突出了从事差异信息处理以产生行为的相互连接节点的作用。许多天生的社会行为对于生殖适应性至关重要,因此在乳腺和复制剂中,在哺乳动物的生产过程中,在哺乳动物的早期发展中,在男性和女性的基本上不同。社会行为及其成人表现形式的早期生活编程是独立的,但却是封锁的,但尚不清楚。本综述旨在通过识别四种核心机制(表观遗传学,细胞死亡,电路形成和成人Hormonal调制)来强调这一差距,从而将发展变化与交配和侵略的成人行为联系起来。我们进一步建议,一种独特的社会行为,青少年的玩法,通过参与基于成人生殖和侵略性行为的相同神经网络来弥合前白大学的大脑。

肝脏在大鼠后HPA轴成熟和性二态性
下丘脑 - 垂体 - 肾上腺(HPA)轴的正常功能对于生存至关重要,并且为年龄,性别和上下文特定的作用编排了其发育。肝脏会影响HPA个体发育,整合了抑制或激活其发育的多种内分泌信号。本综述研究了多种内分泌系统中肝坐标产后变化中基因表达的发育变化如何促进大鼠HPA轴的成熟和性二态性。具体而言,它研究了睾丸雄激素产生的个体发育,生长增生的激素活性和下丘脑 - 垂体 - 甲状腺甲状腺轴的活性如何相交以影响肝蛋白样生长因子1的肝基因表达,类似糖皮质激素结合的球素,甲状腺素固定素 - 11-11- 11- 1和5α-还原酶类型1。这种分子变化的时机在哺乳动物物种之间有所不同,但它们在进化上是保守的,并且有望在广泛地控制稳态,尤其是在逆境中。重要的是,这些多样的内分泌系统以肝脏为Nexus,在整个产后发育过程中建立了HPA轴的基本组织,从而最终确定了成年期糖皮质激素的作用。

与先天免疫中的性二态性和对果蝇果蝇感染的反应
性别在对感染的反应和自身免疫的发展上显示出深远的差异。从节肢动物到脊椎动物,免疫反应中的二态性二态性在整个类群中无处不在。果蝇Melanogaster在基线,致病性挑战和衰老时显示出强烈的性二态性。我们对果蝇免疫的同行评审文献进行了详尽的调查,并介绍了一个出版物数据库,指示每项研究中分析的性别(ES)。尽管我们发现对社区对成人免疫的兴趣日益增加,并且在报告两性时,该领域的主要工作仅使用一种性行为,或者不按性分层进行分层。我们合成了对细菌,病毒和真菌感染的性二态反应的证据。二态性可能是由不同的免疫区室介导的,我们回顾了有关行为,上皮,细胞和全身性(脂肪体介导的)免疫的性别差异的工作。在检查免疫组织,免疫衰老和炎症的性二态衰老方面的新兴工作。我们考虑了免疫投资中性别差异的进化驱动因素,突出了果蝇生物学的特征,这些特征使其特别适合对免疫二态性的研究,并讨论了未来探索的领域。

Pomc 在两性体型二态性中起着重要作用...
© 2021 作者,经 Springer Science+Business Media, LLC 独家授权,隶属于 Springer Nature。这是发表在《海洋生物技术》上的文章的同行评审后、编辑前的版本。最终认证版本可在线获取:http://dx.doi.org/10.1007/s10126-020-10015-2。
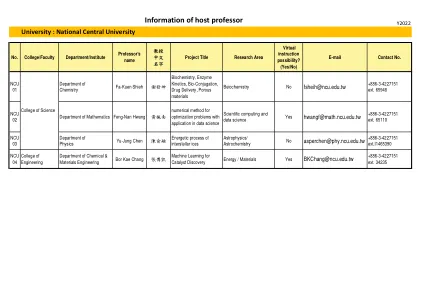
附件二、host professor information form_NCU.xlsx
人工智能(AI),深度学习,智能无人机,智能运输系统(ITS),强化学习(RL),神经建筑搜索(NAS),车辆品牌和模型识别,基于KINECT - 基于Kinect的在线手写识别系统,人员计数,行为分析,车牌检测和识别系统