XiaoMi-AI文件搜索系统
World File Search System成肌细胞

ZBED6‐IGF2 轴对小鼠成肌细胞代谢调节的重要性
摘要转录因子 ZBED6 充当 Igf2 的抑制因子,并直接或间接影响数千个基因的转录调控。在这里,我们在小鼠 C2C12 成肌细胞中使用基因编辑,并表明 ZBED6 仅通过其在 Igf2 内含子 1 中的结合位点 5′-GGCTCG-3′ 来调节 Igf2。删除这个基序(Igf2 ΔGGCT)或完全消融 Zbed6 会导致 IGF2 蛋白上调约 20 倍。定量蛋白质组学显示,在 Zbed6 −/− 和 Igf2 ΔGGCT 成肌细胞中 Ras 信号通路均被激活,并且在 Zbed6 −/− 成肌细胞中表现出表达改变的蛋白质中线粒体膜蛋白显著富集。Zbed6 −/− 和 Igf2 ΔGGCT 成肌细胞均表现出更快的生长速度并发展为肌管肥大。由于 IGF2 上调,这些细胞表现出 O 2 消耗率增加。转录组分析显示,Zbed6 −/− 和 Igf2 ΔGGCT 肌管中差异表达基因的重叠度约为 30%,其中上调的基因与肌肉发育有关。相反,成肌细胞中的 ZBED6 过表达导致细胞凋亡、细胞周期停滞、线粒体活动减少以及成肌细胞分化停止。在 Zbed6 −/− 和 Igf2 ΔGGCT 成肌细胞中观察到的生长和分化表型的相似性表明 ZBED6 主要通过调节 IGF2 表达来影响线粒体活动和肌肉生成。这项研究为 ZBED6-Igf2 轴如何影响肌肉代谢提供了新的见解。

MyoD–Dll1 轴连接的 Notch 信号与肌肉生成的反馈调节
肌肉前体细胞称为成肌细胞,对肌肉发育和再生至关重要。Notch 信号是一种古老的细胞间通讯机制,在控制成肌细胞的成肌程序中起着重要作用。目前,成肌信号是否以及如何反馈来改善这些细胞中的 Notch 活性在很大程度上是未知的。在这里,通过小鼠和人类基因获得/功能丧失研究,我们报告 MyoD 直接开启 Notch 配体基因 Dll1 的表达,该基因激活 Notch 通路以防止邻近成肌细胞的预防性分化,同时自主抑制 Notch 以促进 Dll1 表达细胞中的成肌程序。在机制上,我们通过表征新型 E-box 缺陷小鼠模型的肌肉生成,以及通过 CRISPR 介导的干扰在人类细胞中研究了体内 MyoD-Dll1-Notch 轴背后的顺式调控 DNA 基序。这些结果揭示了介导 Notch 和肌肉生成相互控制的关键转录机制。

Prime编辑允许在负责Duchenne肌肉营养不良的基因中引入特定突变
摘要:从CRISPR/CAS9发现得出的主要编辑技术允许在特定基因中对选定的核苷酸进行修改。我们用它在外显子9、20、35、43、55和61中插入了特定的点突变,该基因肌营养不良蛋白编码为肌营养不良蛋白,该基因在DMD患者中不存在。分别用Prime Editor 2(PE2)和PE3获得了HEK293T细胞中DMD基因的11%和21%所需的突变。三种重复治疗将PE2的特定突变的百分比增加到16%。在单次治疗后,原始的邻接基序(PAM)序列中的额外突变提高了PE3结果至38%。我们还对患者成肌细胞中DMD基因的外显子6中的外显子6中的c.428 g>进行了校正。成肌细胞电穿孔分别显示高达8%和28%的修饰。成肌细胞校正导致通过蛋白质印迹检测到的肌管中肌营养不良蛋白的表达。因此,可以使用序数编辑来校正DMD基因中的点突变。

无动物的生长培养基解决方案用于细胞农业
采购单元是培养肉类生产的工作流程的第一阶段。栽培的肉取决于从动物那里收集细胞。原代成年干细胞是一种选择,是从组织活检或验尸组织获得的。多能细胞来源 - 胚胎干细胞(ESC)或诱导的多能干细胞(IPSC) - 是另一种选择。原代细胞也可以从动物中分离出来。在成年干细胞中,从动物中收集的骨骼肌组织可用于分离肌肉常驻祖细胞。作为胚胎干细胞是多能细胞的,它们可以引起任何非生殖细胞类型。iPSC需要通过区分中胚层细胞谱系以获得肌肉居民祖细胞细胞类型8,将IPSC重新编程回到类似的多能状态。干细胞可以分化为制造肉类的细胞类型,包括成肌细胞/成肌细胞,脂肪细胞(脂肪细胞)和成纤维细胞9。一旦获得了足够数量的细胞,报告说10 12-10 13个细胞10至100 kg肉10,祖细胞可以终止分化为成熟的细胞或组织。
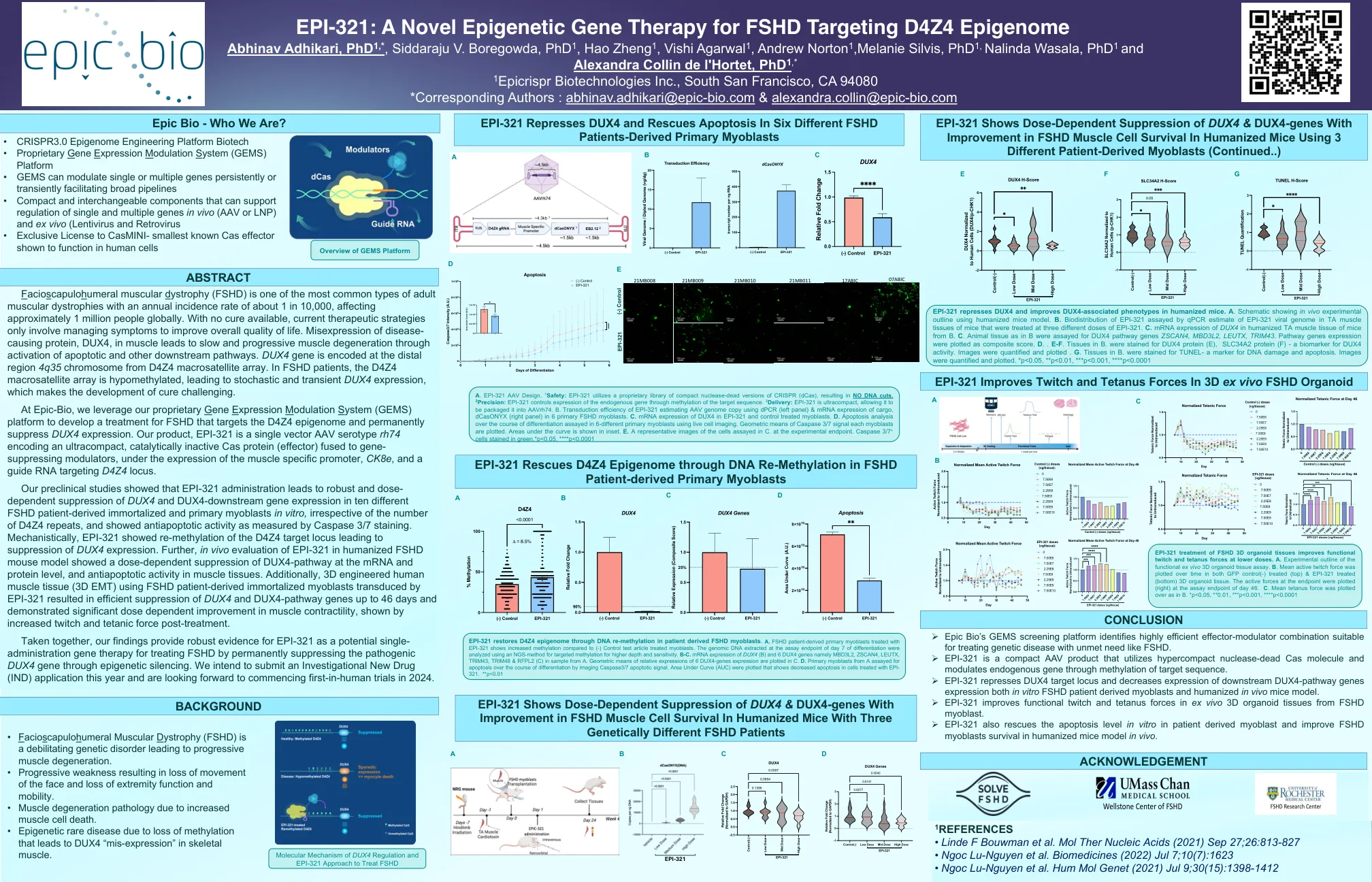
Epi-321-A-Novel-epbepenetic-Gene-gene-for-fshd- ...
我们的临时性研究表明,Epi-321的给药可在十种不同的FSHD患者衍生的患者衍生的永生化和原发性成肌细胞中对Dux4和Dux4-Downstream基因表达的稳健和剂量抑制,无论D4Z4重复序列的数量如何,并且表现出抗iapoptication Asspase 3 Inge caspase 3 ige。从机械上讲,Epi-321显示了D4Z4靶基因座的重新甲基化,从而导致DUX4表达抑制。此外,在人源化FSHD小鼠模型中对Epi-321的体内评估显示,在mRNA和蛋白质水平上对Dux4-Pathway的剂量依赖性抑制作用,以及肌肉组织中的抗凋亡活性。此外,使用FSHD患者衍生的永生化的成肌细胞(Epi-321)进行了3D设计的人体肌肉组织(3D EMT),从而有效地抑制了DUX4和DUX4-PATHWAY基因长达46天,并显示出肌肉缩减性的剂量依赖性,表现出了肌肉缩减的显着改善,表现出受肌肉的增长和Teteratient poptertic posteatial posteat posteat teteat teteat awteat aTteat aTteat aTeat eateat aTeat酸味。

DNA 损伤反应随肌源性分化而下降
DNA 完整性不断面临诱导 DNA 损伤的物质的威胁。所有生物体都配备了 DNA 损伤反应机制网络,可以修复 DNA 损伤并恢复正常的细胞活动。尽管在复制细胞中已经揭示了 DNA 修复机制,但人们对 DNA 损伤在有丝分裂后细胞中的修复方式仍然知之甚少。肌纤维是高度特化的有丝分裂后细胞,以合胞体形式组织,在放射治疗后容易发生与年龄相关的退化和萎缩。我们研究了肌纤维核的 DNA 修复能力,并将其与增殖性成肌细胞中的测量值进行了比较。我们重点研究了纠正电离辐射 (IR) 诱导损伤的 DNA 修复机制,即碱基切除修复、非同源末端连接和同源重组 (HR)。我们发现,在分化程度最高的成肌细胞肌管中,这些 DNA 修复机制表现出 DNA 修复蛋白向 IR 损伤 DNA 募集的动力学减弱。对于碱基切除修复和 HR,这种减弱可能与参与这些过程的关键蛋白的稳态水平降低有关。

牲畜细胞类型具有肌源性分化电势
随着大规模动物生产的当前环境影响以及对农场动物福利的关注,研究人员正在质疑我们是否可以为食品生产而培养动物细胞。本综述着重于细胞农业领域的关键方面:细胞。我们总结了目前用于开发培养肉类的农场动物的各种细胞类型的信息,包括间充质基质细胞,成肌细胞和多能干细胞。审查研究了每种细胞类型的优点和局限性,并考虑了选择适当的细胞来源以及影响细胞性能的细胞培养条件等因素。由于目前在培养的肉类中的研究旨在创建肌肉纤维来模仿肉的质地和营养谱,因此我们专注于细胞的肌源分化能力。用于此目的的最常用的细胞类型是成肌细胞或卫星细胞(SC S),但是鉴于它们的增殖能力有限,正在努力为间充质基质细胞(MSC S)和多能干细胞制定肌生成分化方案(PSC S)。后一种细胞类型的多能特征可能会使肉类中发现的其他组织(例如脂肪和结缔组织)产生。本综述可以帮助指导在培养的肉类发育的背景下选择细胞类型或培养条件。

介导 DMD 基因外显子跳跃的主要编辑策略
杜氏肌营养不良症是一种罕见且致命的遗传性疾病,因 DMD 基因突变导致进行性肌肉萎缩。我们使用 CRISPR-Cas9 Prime 编辑技术开发了不同的策略来纠正 DMD 基因中外显子 52 或外显子 45 至 52 缺失的移码突变。使用优化的 epegRNA,我们能够在高达 32% 的 HEK293T 细胞和 28% 的患者成肌细胞中诱导外显子 53 剪接供体位点的 GT 核苷酸的特异性替换。我们还分别在 HEK293T 细胞和人类成肌细胞中实现了外显子 53 的 GT 剪接位点的 G 核苷酸的缺失高达 44% 和 29%,以及在外显子 51 的 GT 剪接供体位点之间插入 17% 和 5.5% 的 GGG。修改外显子 51 和外显子 53 的剪接供体位点可引发它们的跳跃,从而分别允许外显子 50 连接到外显子 53 和外显子 44 连接到外显子 54。如蛋白质印迹所示,这些修正恢复了肌营养不良蛋白的表达。因此,使用 Prime 编辑在外显子 51 和 53 的剪接供体位点诱导特定的替换、插入和缺失,以分别纠正 DMD 基因中携带外显子 52 和外显子 45 至 52 缺失的移码突变。

果蝇肌原性抑制剂他的基因对于成人肌肉功能和肌肉干细胞维持至关重要
果蝇肌生成抑制剂他的基因是成人肌肉功能和肌肉干细胞维护的essen0al,Robert Mitchell-Gee*1,Robert Hoff*2,Robert Hoff*2,Kumar Vishal2,3,Daniel Hancock1,Daniel Hancock1,Sam McKitrick4,Sam McKitrick4,Cristina newnes newnes newnes-quipperjeta1,tyna and crippation and tyanna l.lovator richana l.lovator,richanna l.lova。 taylor1+ 1。生物科学学院,加的夫大学,加的夫,CF10 3AX,英国。2。圣地亚哥州立大学生物学系,圣地亚哥,加利福尼亚州92182,美国3。圣何塞州立大学生物科学系,圣何塞,加利福尼亚州95192,美国4。 新墨西哥大学的生物学系,美国新墨西哥州87131,美国 *这些作者同样贡献了 +通讯作者:taylormv@cardiff.ac.uk摘要脊椎动物肌肉纤维的群群群群肌肉干细胞(Muscs),或“卫星细胞),或“卫星细胞”,对肌肉的增长,可容纳肌肉,可容纳和修复。 在果蝇中,直到最近才描述了具有相似特征的成年MUSC。 这打开了果蝇系统,用于分析MUSC在肌肉维护,修复和衰老中的运作方式。 在这里,我们表明HIM基因在成年肌肉祖细胞(AMP)或成肌细胞中表达,这使成年果蝇胸腔飞行和跳跃肌肉表达。 值得注意的是,我们还表明,他在飞行肌肉中表达了他,将他识别为这些昆虫MUSC的第二个遗传标记。 然后我们探索了他的功能。 他的突变体破坏了胸跳肌肉的组织,导致跳跃能力降低。 他的突变体还减少了成肌细胞的池,会发展为飞行肌肉。 2015; Laurichesse and Soler 2020)。圣何塞州立大学生物科学系,圣何塞,加利福尼亚州95192,美国4。新墨西哥大学的生物学系,美国新墨西哥州87131,美国 *这些作者同样贡献了 +通讯作者:taylormv@cardiff.ac.uk摘要脊椎动物肌肉纤维的群群群群肌肉干细胞(Muscs),或“卫星细胞),或“卫星细胞”,对肌肉的增长,可容纳肌肉,可容纳和修复。 在果蝇中,直到最近才描述了具有相似特征的成年MUSC。 这打开了果蝇系统,用于分析MUSC在肌肉维护,修复和衰老中的运作方式。 在这里,我们表明HIM基因在成年肌肉祖细胞(AMP)或成肌细胞中表达,这使成年果蝇胸腔飞行和跳跃肌肉表达。 值得注意的是,我们还表明,他在飞行肌肉中表达了他,将他识别为这些昆虫MUSC的第二个遗传标记。 然后我们探索了他的功能。 他的突变体破坏了胸跳肌肉的组织,导致跳跃能力降低。 他的突变体还减少了成肌细胞的池,会发展为飞行肌肉。 2015; Laurichesse and Soler 2020)。新墨西哥大学的生物学系,美国新墨西哥州87131,美国 *这些作者同样贡献了 +通讯作者:taylormv@cardiff.ac.uk摘要脊椎动物肌肉纤维的群群群群肌肉干细胞(Muscs),或“卫星细胞),或“卫星细胞”,对肌肉的增长,可容纳肌肉,可容纳和修复。在果蝇中,直到最近才描述了具有相似特征的成年MUSC。这打开了果蝇系统,用于分析MUSC在肌肉维护,修复和衰老中的运作方式。在这里,我们表明HIM基因在成年肌肉祖细胞(AMP)或成肌细胞中表达,这使成年果蝇胸腔飞行和跳跃肌肉表达。值得注意的是,我们还表明,他在飞行肌肉中表达了他,将他识别为这些昆虫MUSC的第二个遗传标记。然后我们探索了他的功能。他的突变体破坏了胸跳肌肉的组织,导致跳跃能力降低。他的突变体还减少了成肌细胞的池,会发展为飞行肌肉。2015; Laurichesse and Soler 2020)。在飞行肌肉本身中,他的突变体的MUSC数量依赖于年龄,这表明他是维持成年肌肉干细胞种群所必需的。此外,MUSC的这种下降与功能效应相吻合:飞行能力的年龄下降。总的来说,他是果蝇成人MUSC的新颖标志,并且在老化过程中需要保持MUSC数量和飞行能力。介绍。在水果中,果蝇果蝇已证明了研究人员探索肌肉发育的遗传和细胞基础的宝贵模型(Dobi等人在发育过程中,果蝇经历了两波骨骼肌肌发生。胚胎发生过程中的第一个引起了使用ungl pupagon的幼虫肌肉。第二波在普帕奇(Pupagon)期间形成了在成年型中发现的各种肌肉,这些肌肉持续了两个到三个月。不同的成年肌肉是由成年肌肉祖细胞(AMP)引起的,这是一种干细胞populagon,在胚胎发生过程中被放在一边,然后在幼虫寿命中增殖。成年肌肉包括由机翼圆盘AMP形成的胸间间接肌(IFMS)和跳跃肌肉(也称为TDT,TDT,TREGAL的to骨抑制剂),这些肌肉是由与T2间胸乳清盘(Jaramillo et e e2009)。

AAV 载体中新型组合 microRNA 结合位点协同减少抗原呈递和转基因免疫,实现高效、稳定的转导
重组腺相关病毒 (rAAV) 平台有望用于体内基因治疗,但抗原呈递细胞 (APC) 的不良转导会削弱其应用前景,而抗原呈递细胞又会引发宿主对 rAAV 表达的转基因产物的免疫。鉴于最近接受高剂量全身 AAV 载体治疗的患者出现的不良事件,推测这些不良事件与宿主的免疫反应有关,开发抑制先天性和适应性免疫的策略势在必行。使用 miRNA 结合位点 (miR-BS) 来赋予内源性 miRNA 介导的调控,使转基因表达脱离 APC,有望降低转基因免疫力。研究表明,将 miR-142BSs 设计到 rAAV1 载体中能够抑制树突状细胞 (DC) 中的共刺激信号、减弱细胞毒性 T 细胞反应并减弱小鼠转导肌细胞的清除,从而允许在肌纤维中持续转基因表达,同时几乎不产生抗转基因 IgG。在本研究中,我们针对 26 种在 APC 中大量表达但在骨骼肌中不表达的 miRNA 筛选了单个和组合 miR-BS 设计。高免疫原性卵清蛋白 (OVA) 转基因被用作外来抗原的替代物。在成肌细胞、小鼠 DC 和巨噬细胞中进行的体外筛选表明,miR-142BS 和 miR-652-5pBS 的组合强烈抑制了 APC 中的转基因表达,但保持了成肌细胞和肌细胞的高表达。重要的是,携带这种新型 miR-142/652-5pBS 盒的 rAAV1 载体在小鼠肌肉注射后比以前的去靶向设计实现了更高的转基因水平。该盒强烈抑制细胞毒性 CTL 激活和