XiaoMi-AI文件搜索系统
World File Search System蝶啶
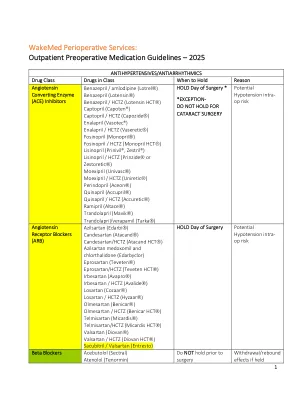
门诊术前用药指南 – 2025 年
依普利酮 (Inspra ® ) 依他尼酸 (Edecrin ) 呋塞米 (Lasix®) 氢氯噻嗪 (Microzide , Esidrix®) 吲达帕胺 (Lozol ) 美托拉宗 (Zaroxolyn ) 甲唑胺 甲氯噻嗪 美托拉宗 (Zaroxoxlyn ) 螺内酯 (Aldactone ) 螺内酯 / 氢氯噻嗪 (Aldactazide ) 托拉塞米 (Demadex ) 氨苯蝶啶 (Dyrenium ) 氨苯蝶啶 / HCTZ (Dyazide , Maxzide ) 他汀类药物 阿托伐他汀 (Lipitor) 氟伐他汀 (Lescol) 洛伐他汀(Mevacor) 匹伐他汀(Livalo) 普伐他汀(Pravachol) 瑞舒伐他汀(Crestor) 辛伐他汀(Zocor)

利用 CRISPR/Cas9 高效修饰玻璃翅神枪手 Homalodisca vitripennis (Germar) 基因组
CRISPR/Cas9 技术可将遗传技术扩展到以前无法进行遗传分析的昆虫害虫。我们报告了在玻璃翅尖足猎蝽 (GWSS) Homalodisca vitripennis 中建立遗传分析的结果,该昆虫是加利福尼亚州农业中一种重要的叶蝉害虫。我们使用一种新颖而简单的方法,在宿主植物上进行原位胚胎显微注射,获得了超过 55% 的朱砂和白色眼色素位点的高频率诱变。通过配对交配,我们获得了 100% 的 w 和 cn 等位基因到 G3 代的传递,并且还确定了这两个基因都位于常染色体上。我们对翅膀表型的分析意外地发现了蝶啶色素参与了翅膀和翅脉着色,表明这些色素的作用不仅限于眼睛颜色。我们利用扩增子测序来检查注射卵对成虫造成的脱靶诱变程度,结果发现脱靶诱变程度可以忽略不计或根本不存在。我们的数据表明,GWSS 可以轻松开发为半翅目昆虫的遗传模型系统,从而能够研究有助于入侵害虫和植物病原体载体成功的特征。这将促进新的遗传控制策略。

引导 RNA 结构设计使组合 CRISPRa 程序能够进行生物合成分析
改造细菌代谢以有效地从多步骤途径产生化学物质和材料需要优化多基因表达程序以实现酶平衡。CRISPR-Cas 转录控制系统正在成为编程多基因表达调控的重要代谢工程工具。然而,向导 RNA 折叠的可预测性较差会通过不可靠的表达控制破坏酶平衡。我们设计了一组可以描述向导 RNA 折叠的计算参数,我们预计它们可以广泛适用于 CRISPR-Cas9 系统。在这里,我们将修饰的向导 RNA (scRNA) 对大肠杆菌中 CRISPR 激活 (CRISPRa) 的功效与描述折叠成活性结构的速率的动力学参数相关联。此参数还支持正向设计新的 scRNA,在我们的筛选中没有观察到失败。我们使用来自该组的 CRISPRa 靶序列来设计一个由三个合成启动子组成的系统,该系统可以在 >35 倍的动态范围内正交激活和调整所选输出的表达。独立的激活调节允许通过 64 个成员的组合三重 scRNA 库对三维表达设计空间进行实验探索。我们将这些 CRISPRa 程序应用于两种生物合成途径,证明了大肠杆菌中有价值的蝶啶和人乳寡糖产品的生产。对这些设计空间进行分析表明,表达组合产生的滴度比最大表达产生的滴度高出 2.3 倍。映射生产还可以确定瓶颈作为途径重新设计的目标,将寡糖乳糖-N-四糖的滴度提高 6 倍。在计算 scRNA 功效预测的帮助下,组合 CRISPRa 策略能够有效优化多步骤代谢途径。更广泛地说,这里揭示的引导 RNA 设计规则可能使有效的多引导程序的常规设计成为可能,用于细菌宿主中 CRISPR 基因调控的广泛模型和数据驱动应用。