XiaoMi-AI文件搜索系统
World File Search System酶系统

Prime editor 整合酶系统促进靶向 DNA 插入及其他
转录模板和引物结合位点(图 1 A) [1]。无需诱导 DSB 和提供 DNA 供体,prime editing 是一种很有前途的精准基因组编辑技术。Prime edit 最初效率较低 [1]。令人鼓舞的是,通过共表达占主导地位的负错配修复 (MMR) 蛋白来抑制 MMR 去除 prime edit 的异源双链 DNA 产物 [2],或通过使用具有多核苷酸替换 [3] 或减少 3′ 端降解 [4] 的 pegRNA,取得了改进。Prime edit 通过所有 12 个碱基替换和不同长度的删除,大大提高了基因组编辑能力 [1,5]。然而,一个挑战是使用 PE 插入超过 40 bp 的长 DNA,这部分是由于 pegRNA 3′延伸的长度限制和较长 RNA 的转录效率降低 [1]。为了克服目前 Prime Editing 对长 DNA 插入的限制,最近的两项研究报道了 Prime Editor 整合酶 (PEI) 系统,该系统无缝结合了 Prime Editing 和基于丝氨酸整合酶的重组 [ 6 , 7 ]。
微生物学重点8.1
使用传统的与改进的含有色彩基质的培养基制剂是当前在微生物学领域中的重要主题。这种进步背后的动力是培养基的生产,这将使微生物的检测和鉴定更加快速,更可靠。发色底物(例如ONPG,X-GAL或X-GLU)以及培养基的指定选择性,是发色介质背后的简单原理。目标生物的特征是特定的酶系统,这些酶系统将底物代谢以释放发色原(见图2)。然后,可以通过直接观察液体汤或琼脂板上的菌落中的明显颜色变化来视觉检测到发色原。有时可以直接确认目标生物,而无需进一步测试。当前,也可以检测和区分同一板上的多个生物体。借助几种发色底物和足够的选择性的组合,可以在一块板上区分几种或基团的微生物。在表1中,已知的底物和选择性代理被列出,并对其他可能性给出一些了解。

醛氧化酶 (AO) 反应表型分析
“AOX 依赖性新药候选物的生物转化是一个新兴问题,因为旨在减少 CYP450 依赖性代谢的化学合成新策略往往会富集药效团,而药效团是 AOX 底物,并被该酶系统灭活。这要求开发新的方法来预测和测试 AOX 依赖性代谢。” 2

3HP 药物间相互作用,包括 ART
当两种药物一起服用时,其中一种药物对身体的作用可能会发生变化。药物相互作用 (DDI) 可能会增加或减少一种或两种药物的作用,或导致不良事件。因此,DDI 可能会延迟、增强或减少同时服用的两种药物的吸收。许多 DDI 是由肝脏中的一种酶系统引起的,称为细胞色素 P450。这些酶可以被诱导,从而更活跃,也可以被抑制,从而活性降低。当活性更高时,它们会增加通过同一系统的其他药物的代谢,并降低这些药物的浓度。这会使第二种药物无效。当酶复合物的活性降低时,所服用的药物的代谢就会降低,体内的浓度就会升高。然后它们会导致更多的不良事件和副作用。

CRISPR / Cas9基因组编辑技术在人类疾病建模和基因治疗中的应用和前景
成簇的规律间隔短回文重复序列 (CRISPR) 介导的 Cas9 核酸酶系统已广泛用于真核细胞中的基因组编辑和基因修饰。CRISPR/Cas9 技术在各种应用中具有巨大潜力,包括纠正人类基因组内的遗传缺陷或突变。CRISPR/Cas9 基因组编辑系统在人类疾病研究中的应用有望解决生命科学研究中遇到的众多复杂的分子生物学挑战。在这里,我们回顾了 CRISPR/Cas9 技术的基本原理及其在神经退行性疾病、心血管疾病、自身免疫相关疾病和癌症中的最新应用,重点关注 CRISPR/Cas9 在这些疾病中的疾病建模和基因治疗潜力。最后,我们概述了使用 CRISPR/Cas9 技术进行疾病研究和治疗的局限性和未来前景。
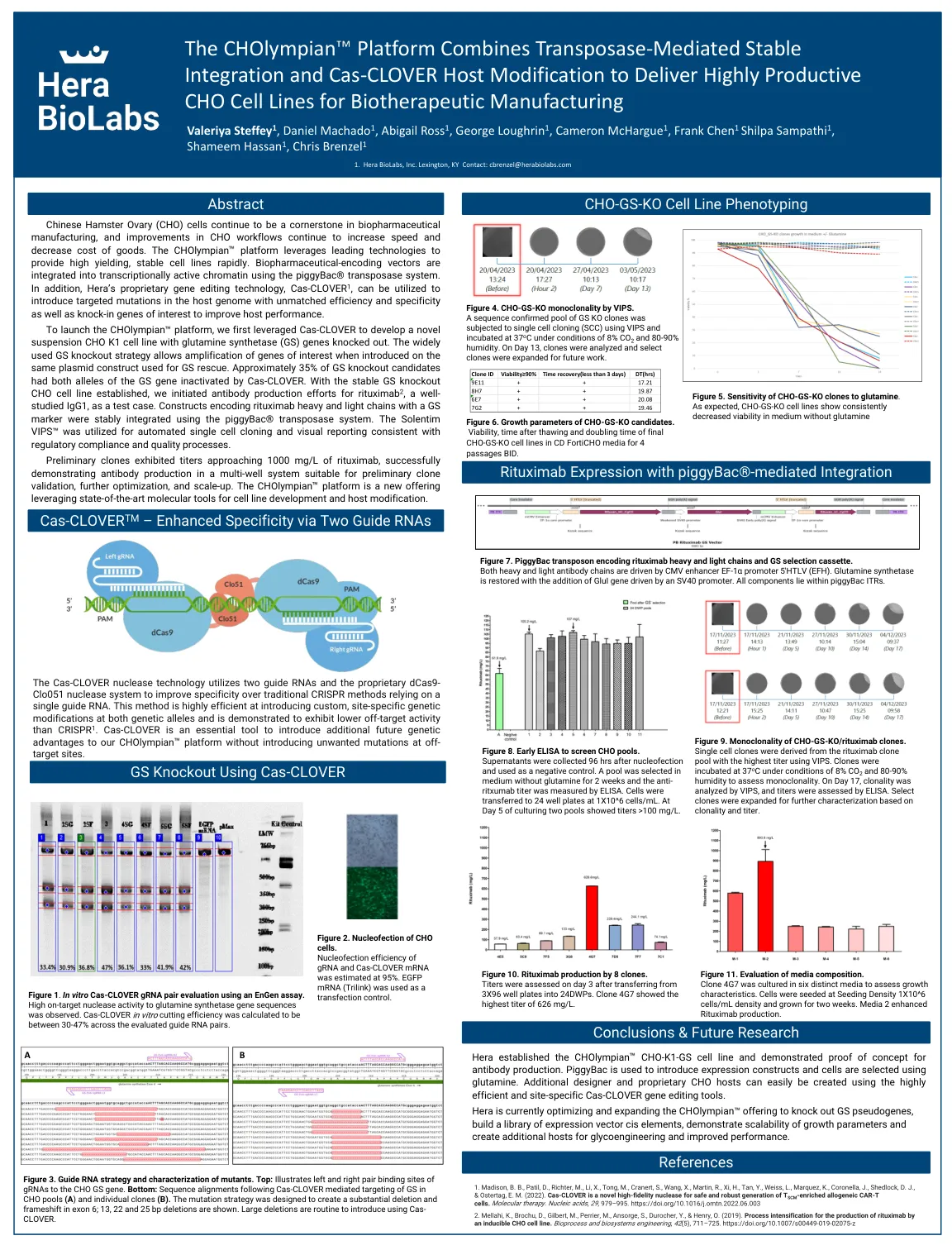
CHOlympian 平台结合转座酶...
平台,我们首先利用 Cas-CLOVER 开发了一种敲除谷氨酰胺合成酶 (GS) 基因的新型悬浮 CHO K1 细胞系。广泛使用的 GS 敲除策略允许在用于 GS 拯救的相同质粒构建体上引入目的基因时对其进行扩增。大约 35% 的 GS 敲除候选细胞的两个等位基因都被 Cas-CLOVER 灭活。在建立稳定的 GS 敲除 CHO 细胞系后,我们开始针对利妥昔单抗 2(一种研究充分的 IgG1)进行抗体生产,作为测试案例。使用 piggyBac® 转座酶系统稳定整合了编码具有 GS 标记的利妥昔单抗重链和轻链的构建体。Solentim VIPS

CRISPR-Cas12a 在多细胞生物中的当前和未来应用
摘要 CRISPR-Cas 系统在原核生物对噬菌体和外来质粒等移动遗传元件的适应性免疫中起着关键作用。在过去十年中,Cas9 已成为一种强大而多功能的基因编辑工具。随之而来的是,新型 RNA 引导的核酸内切酶系统 CRISPR-Cas12a 因其独特的特性(例如其高特异性或靶向富含 T 基序的能力、诱导交错双链断裂和处理 RNA 阵列的能力)正在改变生物学研究。同时,为了实现作物改良、基因治疗、研究模型开发和其他目标,对多细胞生物中高效、安全的基因激活、抑制或编辑的需求日益增加。在本文中,我们回顾了 CRISPR-Cas12a 在多细胞生物中的应用,并讨论了如何克服这些复杂模型特有的挑战,例如外温物种的矢量化或温度变化。

251673G-2BGY20-AT1-SP SHE任务
同种异体供体骨髓移植受到大多数患者的可及性的高度限制,开发新的基因工程技术和程序的社会需求很大,可以使用自体细胞和基因治疗来改善治疗方法。这种治疗方法使用患者来源的诱导多能干细胞(IPSC:具有分化为任何细胞类型的成年重编程细胞)和CRISPR-CAS9设计器核酸酶系统来纠正GLU6VAL突变,以便将其差异化产物用于治疗性移植(Papapapetrou,2017年)。科学作为人类努力的影响方面将在自体细胞和基因替代疗法的使用和接受方面进行探讨,因为它直接受社会和道德考虑的影响。还将检查有关将科学知识应用于治疗的发展和利用的应用和限制方面,由于这种治疗方法中的局限性,这需要相当大的临床前监测。生物背景

DUR董事会会议选秀会议记录2024年6月25日:6:00
ovoquezna®(Vonoprazan)Voquezna®的活性成分Vonoprazan是一种钾富含钾的酸阻滞剂。它通过钾竞争方式抑制H+,K+ -ATPase酶系统,抑制胃顶细胞分泌表面的基础和刺激胃酸分泌。由于该酶被视为顶壁细胞内的酸(质子)泵,因此Vonoprazan被认为是一种胃质子 - 泵抑制剂(PPI),因为它阻止了酸产生的最后一步。vonoprazan不需要通过酸激活。它可以在静止状态和刺激状态下选择性地集中在顶叶细胞中。Vonoprazan以非共价和可逆的方式与活动泵结合。这是为了治愈所有侵蚀性食管炎的愈合以及与成人侵蚀性食管炎相关的胃灼热。维持所有侵蚀性食管炎的愈合以及成人侵蚀性食管炎的胃灼热。与阿莫西林和克拉霉素结合治疗成人幽门螺杆菌(H. Pylori)感染。in

生物修复
微生物烃降解的机制1碳碳氢化合物的降解可以通过特定酶系统(例如氧合酶)介导。2微生物细胞对底物:固定的细胞已用于生物修复多种有毒化学物质。与游离活细胞相比,细菌细胞固定的细胞固定增强了原油的生物降解速率。可以在批处理模式和连续模式下进行固定。包装的床反应器通常在连续模式下用于降解碳氢化合物。3生物生物表面活性剂,这些生物表面活性剂是由多种微生物产生的异质表面活性化学化合物的异质群,例如:假单胞菌sp。4植物修复用于治疗石油烃污染。5基因修饰的细菌:在生物修复中进行基因工程微生物(GEM)的应用已引起了很大的关注,以改善实验室条件下有害废物的降解。因此,微生物降解可以视为石油烃修复的清理策略的关键组成部分。