XiaoMi-AI文件搜索系统
World File Search System双足体

常规足病治疗共付费用为 0 美元。
糖尿病患者需要格外小心和关注双脚。糖尿病会损害双脚的神经末梢和血管,导致双脚受伤时难以察觉,从而导致摔倒。糖尿病还会干扰身体抵抗感染的能力。如果脚部轻微受伤,可能会发展成溃疡或发展成严重感染。

ADM 踝足矫形器和外旋杆的优点
1934 年,伦敦大奥蒙德街医院的丹尼斯·布朗爵士首次描述了传统的靴子和杆式足外展支架,这种支架被国际公认为预防马蹄足复发的标准矫形器。尽管多年来,丹尼斯·布朗支架的概念并没有发生太大的变化,但 C-Pro Direct 最先进的 ADM AFO 和外旋杆代表了重大进步,同时忠实于 Ponseti 博士提出的要求。ADM AFO 和外旋杆的每一个细节都经过精心设计,以最大限度地提高临床表现和患者依从性。该支架更轻、更坚固,外观时尚,同时融入了许多创新设计特点,以促进最佳临床效果。本文档解释了与所有当前替代方案相比,C-Pro Direct 的马蹄足 ADM AFO 和外旋杆马蹄足支架为何以及如何:• 更好地促进伸直外侧边缘的发展并减少高弓足畸形• 更好地促进足部活动性和活动范围的增加• 更好地固定足部,更贴身舒适,比最流行的替代系统轻 32% 且更坚固• 降低皮肤破损、水泡和溃疡的风险• 如果需要更换支架类型,可降低成本• 鼓励更好地遵守支撑协议并获得护理人员/父母的认可• 减少患者在诊所的时间并确保正确应用规定的杆配置• 使马蹄足患者能够从彻底改变现代主流鞋类制造业的先进制造技术中受益最终,这些巨大的优势转化为更好的患者治疗效果和更低的治疗成本。这就是为什么所有马蹄足临床医生现在都应该考虑使用 ADM 模块化支撑系统的原因。

为什么在量子计算中实值振幅就足够了
需要量子计算。对于许多实际问题,仍然需要更快的计算。例如,如果我们能够处理更多数据,目前深度学习的惊人成功(参见 [2])可能会更加惊人。计算机处理信息的能力受到限制,其中一个原因是所有速度都受光速限制。即使以光速,将信号从 30 厘米大小的笔记本电脑的一侧发送到另一侧也需要 1 纳秒 - 在此期间,即使是最便宜的当前计算机也要执行至少 4 次操作。因此,为了加快计算速度,有必要使计算机组件更小。这些组件(例如存储单元)已经由少量分子组成。如果我们将这些细胞做得更小,它们将只由几个分子组成。为了描述如此小物体的行为,有必要考虑量子物理学 - 微观世界的物理学;参见 [1, 4]。因此,计算机需要考虑量子效应。

入侵桡足类基因组适应的进化起源
入侵物种是对生物多样性、生态系统完整性、农业、渔业和公共健康的最大威胁之一,全球每年造成的经济损失高达数千亿美元 1、2。据预测,全球气候变化将以前所未有且复杂的方式增加入侵者的数量和影响 3-8,需要全面了解促进生物入侵成功的机制 9-12。鉴于极小比例的外来物种能够在新栖息地定居,然后成为入侵物种,因此长期以来的争论重点是导致入侵者成功的确切因素 13。人们提出并检验了许多假设,包括繁殖体压力、运输机会、栖息地匹配、繁殖力和种群大小的作用。然而,这些假设并未在不同的分类群和入侵事件中得到一致的实证支持,因此预测能力有限 14-18。 Lee 和 Gelembiuk 19 提出了一种可促进入侵种群出现的进化机制,并假设原生范围内的选择制度是影响入侵成功的关键因素 19 。他们观察到入侵种群往往起源于受到干扰或随时间变化的栖息地 19、20 。因此,他们假设许多入侵种群起源于因环境条件波动而经历平衡选择的原生种群。这种机制往往在相对于环境波动期而言世代时间较短的生物体中起作用,因此不同的等位基因会在不同世代中受到选择的青睐 19 。这种选择制度可以维持原生范围内的遗传变异,并为入侵期间正向选择提供遗传基础 10、15、17、21 – 24 。然而,这一假设此前尚未经过实证检验。平衡选择是自然选择的一种形式,它有利于一个基因座上的多个等位基因,以及它维持地位的能力

'SGLT2抑制剂减轻了狼疮中的足细胞损伤...
我们对Zhao等人的研究充满兴趣和惊讶。对SGLT2抑制剂empagliflozin在全身性红斑狼疮(SLE)和MRLLPR小鼠的狼疮样肾炎中的治疗作用。1关注点是:(1)SGLT2是一种主要在肾脏近端小管中表达的钠葡萄糖转运蛋白。sglt2抑制剂可增强钠和葡萄糖排泄,以及其他机制,这些机制对心脏系统,葡萄糖代谢和造血的有益作用。相比之下,没有直接对自身免疫的影响。作者报告了对SLE的各个方面的抑制以及相关的自动免疫,也就是说,对自动反应性免疫细胞克隆产生的全部IgG和双链DNA(DSDNA)自身抗体的深刻抑制作用,在淋巴机构和骨髓中引起了不可能的效果,这使得对这种疾病的效果不佳,并提高了这种效果。(2)作者试图在人类肾脏活检和MRLLPR小鼠肾脏中的足细胞中降低SGLT2蛋白的表达,但是图2中缺乏管状信号清楚地表明,所使用的抗体未检测到SGLT2。1的确,sglt2在管状细胞的刷子边界中的显着染色,在人类肾脏活检的肾小球中几乎没有表达,抗中性粒细胞胞质抗体(ANCA)血管炎或狼疮2与肾炎2的较低者(scrna)的序列(scrna)不一足细胞中的表达水平。未使用适当的实验工具和控件。这与作者在转基因“ Podocyte”细胞系中发现强SGLT2蛋白表达的发现对比。(3)这种健康和患病的肾脏SCRNA测序数据集中的足细胞对NLRP3转录本也为阴性,因此,关于NLRP3炎性症的参与,所有的数据和推测都与已知的证据无关。尤其是,NLRP3免疫染色(在图4G中)1再次缺乏居民或浸润的单核吞噬细胞中的正信号,识别出所述信号是非特异性的。从这个意义上讲,我们最近反驳了体内原代人足细胞和小鼠足细胞中功能性NLRP3炎症体的主张。3此外,我们在同一小鼠模型中对empagliflozin进行了类似的研究,并且没有观察到任何报告的发现(未提交)。我们认为,狼疮性肾炎患者将在慢性肾脏疾病的进展和相关心血管发病率方面受益于SGLT2抑制作用,但Zhao和SoAthors的报告似乎暗示SGLT2抑制作用将是系统性自动自动抑制的有效抑制器。纸张,就其文章而言,得出的结论不受提供的数据的支持。

利用 CRISPR-Cas9 创建白化鱿鱼品系及其在神经活动体内功能成像中的应用
头足类动物在无脊椎动物中以认知能力、适应性伪装、新颖结构和通过 RNA 编辑重新编码蛋白质的倾向而引人注目。然而,由于缺乏遗传上可处理的头足类模型,这些创新背后的机制尚不清楚。CRISPR-Cas9 等基因组编辑工具允许在不同物种中进行定向突变,以更好地将基因和功能联系起来。一种新兴的头足类模型 Euprymna berryi 产生大量胚胎,这些胚胎可以在其整个生命周期中轻松饲养,并且具有已测序的基因组。作为原理证明,我们在 E. berryi 中使用 CRISPR-Cas9 来靶向色氨酸 2,3 双加氧酶 (TDO) 基因,色氨酸 2,3 双加氧酶 (TDO) 是形成色素色素所需的酶,色素色素是头足类动物眼睛和色素细胞中的色素。将靶向 tdo 的 CRISPR-Cas9 核糖核蛋白注射到早期胚胎中,然后培养至成年。出乎意料的是,注射的标本是有色的,尽管通过对注射动物 (G0s) 进行测序验证了目标位点的插入缺失。经过多代繁殖的 TDO 纯合敲除系也有色。令人惊讶的是,E. berryi 中也存在编码吲哚胺 2,3 双加氧酶 (IDO) 的基因,该酶在脊椎动物中催化与 TDO 相同的反应。使用 CRISPR-Cas9 对 tdo 和 ido 进行双敲除产生了白化表型。我们展示了这些白化病在双光子显微镜对大脑中的 Ca 2+ 信号进行体内成像中的实用性。这些数据表明,制造基因敲除头足类动物系的可行性,可用于对这些行为复杂的生物体的神经活动进行实时成像。

2019财年专利申请技术趋势调查(航天器)
中国空间技术研究院 (中国) 643 26,135 30 空客 (欧洲) 611 13,954 67 波音 (美国) 430 14,624 88 Energiya (俄罗斯) 430 7,401 37 三菱电机 279 89,137 20 IHI 201 13,657 28 泰雷兹 (欧洲) 153 6,495 54 三菱重工 131 27,823 16 霍尼韦尔 (美国) 117 19,431 7 雷神 (美国) 105 5,383 3 斯奈克玛 (欧洲) 102 4,363 6 太空系统/劳拉 (美国) 58 168 12 Viasat (美国) 1 685 0 蓝色起源 (美国) 12 19 1 SpaceX(美国) 1 10 9 Rocket Lab(美国) 5 5 0 北京零度空间科技公司(中国) 2 24 0 Mojave Aerospace Ventures(美国) 2 2 0 PLD space(西班牙) 0 0 0 Reaction Engines(英国) 6 13 4 Relativity Space(美国) 0 2 0 Skyrora(英国) 0 0 0 Oneweb(美国) 11 29 0 Blacksky(美国) 0 0 0 Capella Space(美国) 0 0 0 Hawkeye360(美国) 0 6 0 Iceye(芬兰) 0 1 0 OHB System(德国) 1 8 20 Planet(美国) 5 27 2 Spire Global(美国) 6 22 0 ispace(日本) 7 13 1 Planetary Resources(美国) 4 4 1 Astroscale 12 12 0 D-Orbit (意大利) 4 4 0 NASA (美国) 91 1,924 959 日本宇宙航空研究开发机构 119 500 473 国防科技大学 (中国) 69 6,274 280 哈尔滨工业大学 (中国) 338 25,237 274 加州理工学院 (美国) 19 2,648 314 韩国航空宇宙研究院 (韩国) 436 2,739 72

核融合反应堆高温超导导体开发的现状
名启博:プラマ・核融合学志92,396(2016)。[4 W.H.fietz and al。,IEEE Trans。苹果。超级。26,4800705(2016)。 [5]P。Bruzzone和Al。 ,ncle。 Fuance 58,103001(2018)。 l。米切尔和阿尔。 ,超级条件。 SCI。 树。 34,103001(2021)。 !t。安多和al。 ,技术完整。 1,791(1998)。 Lage F. Dahlgren和Al。 ,Eng已满。 甲板。 167,139(2006)。 ]H。H. Hashizume和Al。 ,Eng已满。 甲板。 63,449(2002)。 [10! Y. Ogawa和Al。 ,J。 填充完整的等离子体。 79,643(2003)。 <+11 Z. Yoshida和Al。 ,Ressing主题等离子体。 1,8(2006)。 [12 Y. Ogawa和Al。 ,Ressing主题等离子体。 9,140,014(2014)。 13 V. Corat和Al。 ,Eng已满。 甲板。 136,1597(2018)。 14 A. Sagara和Al。 ,Eng已满。 甲板。 89,2114(2014)。 15 Y. Zhai和Al。 ,Eng已满。 甲板。 135,324(2018)。 https://typeoneergy.com/ [20! Sorbon和Al。 ,Eng已满。 甲板。 100,378(2015)。 [22 A A. Sykes和Al。26,4800705(2016)。[5]P。Bruzzone和Al。,ncle。Fuance 58,103001(2018)。l。米切尔和阿尔。,超级条件。SCI。 树。 34,103001(2021)。 !t。安多和al。 ,技术完整。 1,791(1998)。 Lage F. Dahlgren和Al。 ,Eng已满。 甲板。 167,139(2006)。 ]H。H. Hashizume和Al。 ,Eng已满。 甲板。 63,449(2002)。 [10! Y. Ogawa和Al。 ,J。 填充完整的等离子体。 79,643(2003)。 <+11 Z. Yoshida和Al。 ,Ressing主题等离子体。 1,8(2006)。 [12 Y. Ogawa和Al。 ,Ressing主题等离子体。 9,140,014(2014)。 13 V. Corat和Al。 ,Eng已满。 甲板。 136,1597(2018)。 14 A. Sagara和Al。 ,Eng已满。 甲板。 89,2114(2014)。 15 Y. Zhai和Al。 ,Eng已满。 甲板。 135,324(2018)。 https://typeoneergy.com/ [20! Sorbon和Al。 ,Eng已满。 甲板。 100,378(2015)。 [22 A A. Sykes和Al。SCI。树。 34,103001(2021)。 !t。安多和al。 ,技术完整。 1,791(1998)。 Lage F. Dahlgren和Al。 ,Eng已满。 甲板。 167,139(2006)。 ]H。H. Hashizume和Al。 ,Eng已满。 甲板。 63,449(2002)。 [10! Y. Ogawa和Al。 ,J。 填充完整的等离子体。 79,643(2003)。 <+11 Z. Yoshida和Al。 ,Ressing主题等离子体。 1,8(2006)。 [12 Y. Ogawa和Al。 ,Ressing主题等离子体。 9,140,014(2014)。 13 V. Corat和Al。 ,Eng已满。 甲板。 136,1597(2018)。 14 A. Sagara和Al。 ,Eng已满。 甲板。 89,2114(2014)。 15 Y. Zhai和Al。 ,Eng已满。 甲板。 135,324(2018)。 https://typeoneergy.com/ [20! Sorbon和Al。 ,Eng已满。 甲板。 100,378(2015)。 [22 A A. Sykes和Al。树。34,103001(2021)。!t。安多和al。,技术完整。1,791(1998)。Lage F. Dahlgren和Al。,Eng已满。甲板。167,139(2006)。]H。H. Hashizume和Al。,Eng已满。甲板。63,449(2002)。[10! Y. Ogawa和Al。,J。填充完整的等离子体。79,643(2003)。<+11 Z. Yoshida和Al。,Ressing主题等离子体。1,8(2006)。[12 Y. Ogawa和Al。,Ressing主题等离子体。9,140,014(2014)。13 V. Corat和Al。,Eng已满。甲板。136,1597(2018)。14 A. Sagara和Al。 ,Eng已满。 甲板。 89,2114(2014)。 15 Y. Zhai和Al。 ,Eng已满。 甲板。 135,324(2018)。 https://typeoneergy.com/ [20! Sorbon和Al。 ,Eng已满。 甲板。 100,378(2015)。 [22 A A. Sykes和Al。14 A. Sagara和Al。,Eng已满。甲板。89,2114(2014)。 15 Y. Zhai和Al。 ,Eng已满。 甲板。 135,324(2018)。 https://typeoneergy.com/ [20! Sorbon和Al。 ,Eng已满。 甲板。 100,378(2015)。 [22 A A. Sykes和Al。89,2114(2014)。15 Y. Zhai和Al。 ,Eng已满。 甲板。 135,324(2018)。 https://typeoneergy.com/ [20! Sorbon和Al。 ,Eng已满。 甲板。 100,378(2015)。 [22 A A. Sykes和Al。15 Y. Zhai和Al。,Eng已满。甲板。135,324(2018)。https://typeoneergy.com/ [20!Sorbon和Al。,Eng已满。甲板。100,378(2015)。[22 A A. Sykes和Al。,ncle。Fusion 58,016039(2018)。<3- y。歌曲和Al。 ,Eng已满。 甲板。 183,113247(2022)。 24-24 N. Yanagi和Al。 ,Ressing主题等离子体。 9,140,013(2014)。 ,Proc。 14th Symp。 Fusion Technology,1727(1986)。歌曲和Al。,Eng已满。甲板。183,113247(2022)。24-24 N. Yanagi和Al。 ,Ressing主题等离子体。 9,140,013(2014)。 ,Proc。 14th Symp。 Fusion Technology,1727(1986)。24-24 N. Yanagi和Al。,Ressing主题等离子体。9,140,013(2014)。,Proc。 14th Symp。 Fusion Technology,1727(1986)。,Proc。14th Symp。Fusion Technology,1727(1986)。

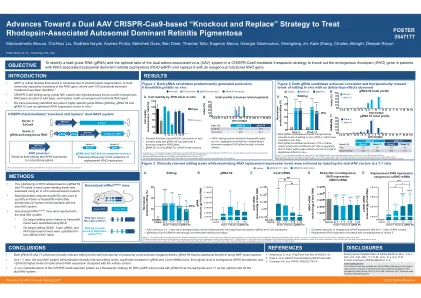
基于双 AAV CRISPR-Cas9 的“...
mRho hRHO/+ 小鼠注射了双 AAV 系统,其中以不同的载体 1:载体 2 比例识别出领先的 gRNA 59,并在注射后 6 周进行分析(载体 n=12;gRNA 59 n=20–22)。显示平均值 (SD)。*p<0.05、**p<0.01、***p<0.001、****p<0.0001 vs 载体。# p<0.05、## p<0.01、#### p<0.0001 vs 其它载体比例。(A) 编辑标准化为转导区域。黑色虚线表示达到治疗相关编辑水平 (≥25%) 的阈值。3 (B) gRNA 水平。(C) Cas9 mRNA 水平。(D) 内源性 hRHO mRNA 水平。数据标签表示与载体相比的百分比下降。(E) 外源性替代 coRHO mRNA 水平。数据标签表示与载体相比的倍数增加。AAV,腺相关病毒;bp,碱基对;coRHO,密码子优化的RHO等位基因;gRNA,向导RNA;hRHO,人类RHO等位基因;mRho,小鼠Rho等位基因;NGS,下一代测序;RHO/Rho,视紫红质;SD,标准差。