XiaoMi-AI文件搜索系统
World File Search System复制


CRISPR复制
摘要 CRISPR/Cas 系统已成为代谢工程和人类基因治疗中基因组编辑的有力工具。然而,使用 CRISPR/Cas 系统在染色体上定位整合异源基因的最佳位点仍是一个悬而未决的问题。选择合适的基因整合位点需要考虑多个复杂的标准,包括与 CRISPR/Cas 介导的整合、遗传稳定性和基因表达相关的因素。因此,在特定或不同的染色体位置上识别此类位点通常需要大量的表征工作。为了应对这些挑战,我们开发了 CRISPR-COPIES,这是一种用于识别 CRISPR/Cas 促进的整合位点的计算流程。该工具利用 ScaNN,这是一种基于嵌入的最近邻搜索的先进模型,可快速准确地进行脱靶搜索,并可在几分钟内识别大多数细菌和真菌基因组的全基因组基因间位点。作为概念验证,我们利用 CRISPR-COPIES 表征了三种不同物种中的中性整合位点:酿酒酵母、Cupriavidus necator 和人类细胞系。此外,我们还为 CRISPR-COPIES 开发了一个用户友好的 Web 界面 (https://biofoundry.web.illinois.edu/copies/)。我们预计 CRISPR-COPIES 将成为靶向 DNA 整合的宝贵工具,并有助于表征合成生物学工具包,实现快速菌株构建以生产有价值的生化物质,并支持人类基因和细胞治疗应用。



DNA-1220-BF26_370W_DATASHEET复制
我们的DNA拆分系列具有令人印象深刻的高级技术,以最大程度地提高性能。双方后侧发电量产生多达30%的能量。我们的专利DNA,双纳米吸收器,技术允许面板从各个方向收集能量,并在高温温度下进行异常运行。

eesl_december_newsletter_301224复制
可再生能源供应链已经变得越来越复杂,这主要是由于与项目组件中使用的原材料有关的问题。例如,据估计,到2030年,稀土金属(如霓虹灯和praseodymium)将缺乏50-60%的短缺,这些金属将用于为风力涡轮机发电机和电动汽车制造高功率磁铁所必需的。太阳能安装需要从多个国家采购的组件。这可能会在运输,关税和遵守跨境运动以及遵守安全,环境和劳动法的情况下造成实际问题。货物的延误可能导致管理库存,项目成本升级和较长的交货时间的困难。包括铁路货运运动在内的多模式运输系统可以为这些挑战提供可靠,可持续和成本效益的解决方案。

最终 - 复制
1美国加利福尼亚大学洛杉矶分校的电气工程系90095,美国2材料科学与工程系,加利福尼亚大学加利福尼亚州洛杉矶分校,加利福尼亚州90095,美国3美国3号电子和X射线仪器,加利福尼亚大学欧文分校,加利福尼亚州Irvine Universition,USAKIAN IRVINA,US 42697,US 42697,USICIAN ELECTIRAILIA,CARICEFERIAN,CARICEFERIAN,USYRIAN,UIVENIA,UIVERIA,>1美国加利福尼亚大学洛杉矶分校的电气工程系90095,美国2材料科学与工程系,加利福尼亚大学加利福尼亚州洛杉矶分校,加利福尼亚州90095,美国3美国3号电子和X射线仪器,加利福尼亚大学欧文分校,加利福尼亚州Irvine Universition,USAKIAN IRVINA,US 42697,US 42697,USICIAN ELECTIRAILIA,CARICEFERIAN,CARICEFERIAN,USYRIAN,UIVENIA,UIVERIA,>


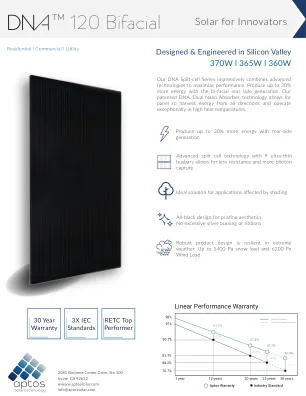
DNA-1220-BF26_370W_DATASHEET复制
我们的DNA拆分系列具有令人印象深刻的高级技术,以最大程度地提高性能。双方后侧发电量产生多达30%的能量。我们的专利DNA,双纳米吸收器,技术允许面板从各个方向收集能量,并在高温温度下进行异常运行。