XiaoMi-AI文件搜索系统
World File Search System多粒

生化技术 26 重组 DNA 技术
克隆载体是一种能够在宿主生物体内复制的 DNA 分子。将目标 DNA 引入该载体以产生重组 DNA 分子。大肠杆菌作为宿主生物,使用多种克隆载体。有时,需要使用不同的宿主进行克隆实验。因此,已经开发了基于其他细菌(如芽孢杆菌、假单胞菌、农杆菌等)和不同真核生物(如酵母和其他真菌)的各种克隆载体。不同类型的 DNA 分子可用作克隆载体,例如它们可能是质粒、噬菌体、粘粒、噬菌粒或人工染色体

高蛋白液体饮食计划 - 手术后前 7-10 天
食用前滤干。• 加 1 勺无味蛋白粉的肉汤 • 牛奶:无脂或 1% • 豆浆:原味或香草味 • Lactaid 牛奶:无脂或 1% • 柔滑口味淡酸奶:低脂(无果粒,用代糖增甜) • 酸奶:原味(无味),低脂 • 希腊酸奶:原味或柔滑口味酸奶,(无果粒),低脂,用代糖增甜 • 自制布丁:用低脂牛奶制成,无糖或不添加糖(可使用无糖布丁粉)
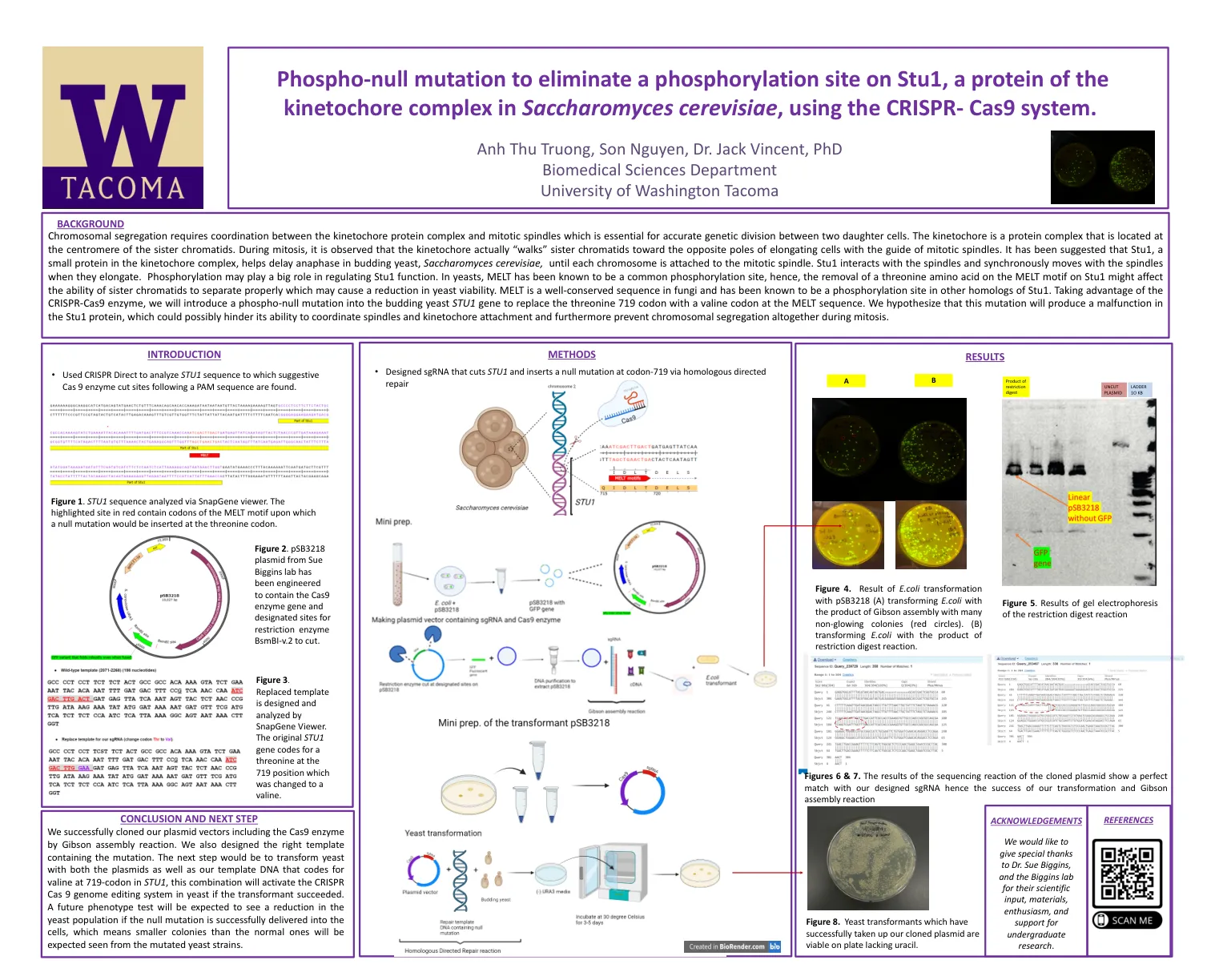
利用 CRISPR 技术进行磷酸化无效突变,消除酿酒酵母动粒复合体蛋白 Stu1 上的磷酸化位点
染色体分离需要动粒蛋白复合物和有丝分裂纺锤体的协调,这对于两个子细胞之间的准确遗传分裂至关重要。动粒是一种位于姊妹染色单体着丝粒的蛋白复合物。在有丝分裂过程中,可以观察到动粒实际上是在有丝分裂纺锤体的引导下将姊妹染色单体“引导”到伸长细胞的相反极点。有人提出,动粒复合物中的小蛋白 Stu1 有助于延迟芽殖酵母酿酒酵母的后期,直到每条染色体都附着在有丝分裂纺锤体上。Stu1 与纺锤体相互作用,并在纺锤体伸长时与其同步移动。磷酸化可能在调节 Stu1 功能方面发挥重要作用。在酵母中,MELT 是一种常见的磷酸化位点,因此,去除 Stu1 上 MELT 基序上的苏氨酸氨基酸可能会影响姐妹染色单体正确分离的能力,从而导致酵母活力下降。MELT 是真菌中保存良好的序列,并且已知是 Stu1 其他同源物中的磷酸化位点。利用 CRISPR-Cas9 酶,我们将在芽殖酵母 STU1 基因中引入磷酸化无效突变,以将 MELT 序列中的苏氨酸 719 密码子替换为缬氨酸密码子。我们假设这种突变会导致 Stu1 蛋白发生故障,这可能会阻碍其协调纺锤体和着丝粒附着的能力,并进一步阻止有丝分裂期间染色体分离。

P1噬菌体实现CRISPR-Cas9对福氏志贺氏菌的抗菌活性
摘要:成簇的、规则间隔的、短回文重复序列 (CRISPR) 和 Cas9 RNA 引导核酸酶的发现为选择性杀死特定种群或物种的细菌提供了前所未有的机会。然而,由于 cas 9 基因构建体无法高效地递送到细菌细胞中,因此 CRISPR-Cas9 在体内清除细菌感染的应用受到了阻碍。在这里,我们使用广宿主范围的 P1 衍生噬菌粒将 CRISPR-Cas9 染色体靶向系统递送到大肠杆菌和引起痢疾的福氏志贺氏菌中,以实现对目标细菌细胞的 DNA 序列特异性杀死。我们表明,辅助 P1 噬菌体 DNA 包装位点 (pac) 的基因改造可显著提高包装噬菌粒的纯度,并改善 Cas9 介导的福氏志贺氏菌细胞的杀灭作用。我们进一步证明,P1 噬菌体颗粒可以使用斑马鱼幼虫感染模型将染色体靶向 cas9 噬菌粒递送到 S. flexneri 体内,从而显著减少细菌负荷并促进宿主存活。我们的研究强调了将基于 P1 噬菌体的递送与 CRISPR 染色体靶向系统相结合以实现 DNA 序列特异性细胞致死率和有效清除细菌感染的潜力。关键词:福氏志贺氏菌、P1 噬菌体、CRISPR-Cas9、抗菌、噬菌粒 ■ 简介


用于分析流体流动中颗粒行为的更简单的粗颗粒模型的开发
摘要:为减少计算量,本文提出了一种新的、更简单的粗粒模型 (SCG),利用离散元法 (DEM) 分析稀薄系统中流体流动下的颗粒行为。在 SCG 模型中,粗粒 (CG) 颗粒以与现有粗粒模型相同的方式从原始颗粒中放大;但是,建模概念与其他模型不同。SCG 模型侧重于流体阻力引起的加速度,CG 颗粒的加速度与原始颗粒的加速度一致。因此,该模型仅施加以下简单规则:颗粒密度与颗粒直径平方的乘积为常数。因此,该模型具有可在 DEM 模拟中轻松实现的功能,以理解建模的物理现象。通过比较均匀和涡流场中 CG 颗粒和原始颗粒的行为来验证该模型。此外,通过在分类器中表示粒子行为,确认了 SCG 模型在模拟真实稀释系统中的可用性。因此,使用 SCG 模型可以更简单地分析稀释粒子浓度系统中的粒子行为。
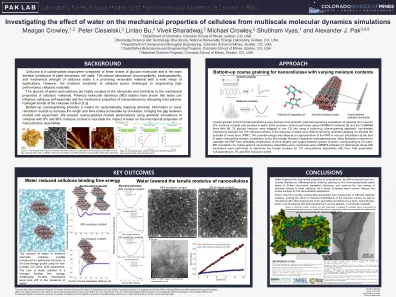
研究水对多尺度分子动力学模拟纤维素机械性能的影响
粗粒(CG)力场参数是使用真空中纤维素Iβ的原子分子动力学模拟得出的(0%的水分含量),并使用Gromacs软件[5]和CHARMM力场进行的水(95%水分含量)溶剂(95%的水分含量)[6]。72使用自下而上的粗粒方法将葡萄糖残基映射到一个CG位置:在存在水存在下,使用雨伞采样确定了100个纤维素表面之间的非键相互作用,以计算平均力的潜力(PMF)。势能被视为真空模拟中PMF的近似值,因为缺乏水减少了对自由能的熵贡献。使用Boltzmann倒置参数化键合的相互作用,以从与CG位点相对应的原子组之间的键长和角度的概率分布来计算PMF。使用LAMMPS软件进行了粗粒纳米纤维素组件的MD模拟[7]。进行了机械应力MD模拟,以确定具有强力场参数的CG纳米纤维素组件的拉伸模量,其水分含量为0%和95%。

阿扎胞苷和 MEK 抑制剂联合治疗 PTPN11 突变的幼年型粒单核细胞白血病的潜在临床应用
幼年型粒单核细胞白血病 (JMML) 是一种罕见的儿童骨髓增生性肿瘤。JMML 的分子特征是 Ras/MAPK 通路过度活跃,最常见的原因是编码蛋白酪氨酸磷酸酶 SHP2 的基因 PTPN11 发生突变。目前治疗 JMML 的策略包括使用低甲基化剂 5-阿扎胞苷 (5-Aza) 或 MEK 抑制剂曲美替尼和 PD0325901 (PD-901),但这些药物均不能作为单一疗法治愈。利用 Shp2 E76K/ + 小鼠 JMML 模型,我们表明 5-Aza 和 PD-901 的组合可调节 JMML 患者中常见的几种血液学异常,部分原因是通过减少白血病造血干细胞和祖细胞 (HSC/Ps) 的负担。接受药物治疗的小鼠中 JMML 特征的减少与接受 5-Aza 和 PD-901 联合治疗的 Shp2 E76K/+ 小鼠中 p-MEK 和 p-ERK 水平的降低有关。RNA 测序分析显示几种 RAS 和 MAPK 信号相关基因减少。此外,在接受两种药物联合治疗的 Shp2 E76K/+ 小鼠中还观察到与炎症和髓系白血病相关的基因表达减少。最后,我们报告了两例接受 5-Aza、曲美替尼和化疗治疗的 JMML 和 PTPN11 突变患者,他们因联合治疗而出现临床反应。

蛋白质Pegylation方案 - 全马提尼粗粒v2.0方案(马提尼,疯狂,gromacs模拟)
-EF =弹性力常数,用于保留蛋白质二级和三级结构。应使用它来测试正常的所有原子构象,以保持所有原子和粗粒结构之间的相似性。应该测试几个值,并且必须使用研究信息来为您的系统选择更好的值。-EL =弹性下键切断。作为-ef标志,必须与晶体学结构进行测试或比较。-EU =弹性上键切断。作为-ef标志,必须与晶体学结构进行测试或比较。-pf =位置约束。用于避免原子运动以平衡系统。应与-p标志一起使用,以选择要约束哪种珠子。骨干是最常见的选择。- 突变=突变一个残基到另一个残基。一般而言,马提尼岛在识别他的HSD:HSD时始终构成始终突变的马提尼岛有一些问题。

交联部分,硬化功能和拓扑质量对热固性形状记忆聚合物的应力恢复的影响:粗粒分子动力学研究
摘要我们通过粗粒分子动力学模拟分析了每个硬汉形状记忆聚合物(TSMP)的交联部分和每个硬汉形状记忆聚合物(TSMP)的应力恢复和拓扑的功能位点数量的影响。通过操纵与每个硬质反应的独特环氧树脂的数量来系统地改变交联网络的质量后,我们发现两种指纹与TSMPS的应力恢复非常相关。这些指纹是连接到两个不同硬化分子的环氧分子的比例,是系统中最大或主要网络的一部分的分子的比例。他们的产品可以用作拓扑评分(S TOPO)来量化网络的拓扑特征。在分析应力恢复与S TOPO的函数时,我们发现S TOPO与恢复应力之间存在很强的相关性。此外,我们观察到,尽管较高的交联部分确实会导致更高的压力恢复,但仍然存在许多例外。高功能硬化剂在相似的S topo处倾向于表现出更高的应力恢复,尤其是在高(> 0.65)的topo处。这些结果表明,增加每个硬化分子的功能位点的数量,结合使用诸如半批量单体添加的方法改善网络拓扑结合,可以改善TSMPS的应力恢复。