XiaoMi-AI文件搜索系统
World File Search System干细胞

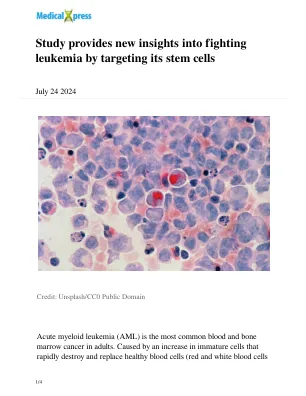
研究通过靶向其干细胞
“使用高级生物信息学技术并与拥抱肿瘤学和精确肿瘤学系的Petros Tsantoulis博士团队合作,我们首先确定这些静态细胞包含由35个基因组成的独特遗传标志。当我们在AML患者的大型临床数据库中使用了这种签名时,我们能够证明该签名与疾病的预后密切相关。”领导这项研究的拥抱肿瘤学


成体干细胞的研究进展及设想
因素的限制。对胚胎干细胞的研究主要是通过动物 实验进行的 , 而成体干细胞 (adult stem cells) 存在 于胎儿和成人各种组织及器官中 , 来源广泛 , 而且不 涉及伦理问题。虽然胚胎干细胞更具有全能性 , 理 论上可生成任何组织 , 容易分化为一些组织如心脏
GCSE干细胞和克隆|活动工作表 GCSE干细胞和分化|考试工作表 GCSE干细胞和克隆|问题
2。为什么患者的身体接受这些干细胞创建的器官?………………………………………………………………………………………………………………………………………………指示每个语句是正确还是错误。i。治疗克隆的过程没有转移病毒感染的风险。

从重组蛋白到干细胞和类器官| ...从重组蛋白到干细胞和类器官| ...
摘要近年来生物制剂在各种疾病中的使用已大大增加。中风是一种脑血管疾病,是第二大最常见的死亡原因,也是全球发病率高的残疾原因。用于用于治疗急性缺血性中风的生物制剂,Alteplase是唯一的溶栓剂。同时,当前的临床试验表明,两种重组蛋白,Tenecteplase和非免疫原性葡萄球菌酶,作为用于急性缺血性中风治疗的新溶栓剂的最有前途的。此外,使用干细胞或类器官进行中风治疗的基于干细胞的治疗在临床前和早期临床研究中显示出令人鼓舞的结果。这些急性缺血性中风的策略主要依赖于未分化的细胞的独特特性来促进组织修复和再生。但是,在这些方法成为常规临床用途之前,仍有一段巨大的旅程。这包括优化细胞输送方法,确定理想的细胞类型和剂量以及解决长期安全问题。本综述介绍了缺血性中风中溶栓治疗的当前或有希望的重组蛋白,并突出了中风治疗中干细胞和大脑器官的前景和挑战。

揭示了干细胞疗法的承诺
摘要:干细胞以其独特的再生能力而闻名,在治疗中风方面具有巨大的希望,这是全球残疾的主要原因。本综述对中风(缺血和出血)恢复中的干细胞应用进行了详细的分析。IT检查基于自体(患者衍生),同种异体(供体)和基于粒细胞的固定刺激因子(G-CSF)干细胞的疗法,重点介绍了细胞类型,例如间质干细胞(MSC),骨莫罗型单核干细胞(骨骼核细胞),骨骼含量单核干细胞(BMMSCSSSC)和NEARARAN和NEARARARARAN/NEARARAN/NEARARAN/NEARARAN/NEARARAN/NEARARITITIRERITITIRERITITOR。本文编译了临床试验数据以评估其有效性和安全性,并解决了这些创新治疗的道德问题。通过解释干细胞诱导的神经系统修复的机制,该评论强调了干细胞在革命中风康复方面的潜力,并提出了未来研究的途径。关键词:干细胞疗法,中风,脑出血,自体干细胞移植,同种异体干细胞移植,粒细胞 - 粘液固体刺激因子

干细胞研究-DEF -LAB
独特的干细胞线标识符BCHI007-A(HNDS0005-01 #B)BCHI007-A-1(HNDS0005-01#B2 + / +)BCHI009-A(HNDS0002-01 #D) BCHi011-A-1 (HNDS0003-01 #F CC39 + / + ) Alternative name(s) of stem cell lines HNDS0005-01 #B HNDS0005-01 #B2 + / + HNDS0002-01 #D HNDS0002-01 #D CC26 + / + HNDS0003-01 #F HNDS0003-01 #F CC39 + / + Institution波士顿儿童医院的联系信息wardiya.afsharsaber@childrens.harvard.edu; Mustafa.sahin@childrens.harvard.edu类型的细胞系IPSCS来源的人类ESC或IPSC BCHI007-A(HNDS0005-01 #B)所需的其他起源信息,年龄:21,性别:性别:f,f,f,种族:白人:白色; BCHI009-A(HNDS0002-01 #D),年龄:20,性别:M,种族:白色; BCHI011-A(HNDS0003-01 #F),年龄:4,性别:M,种族:白色。细胞源成纤维细胞克隆性克隆细胞方法的重编程仙台病毒,非整合(OCT4,SOX2,KLF4和HC-MYC)遗传修饰是

克隆、基因改造和干细胞技术
GSK 的生物伦理和行为准则要求遵守所有外部法律、法规和指南,这些法律、法规和指南适用于将克隆、基因改造和干细胞技术应用于药物研发。以下框架适用于我们代表 GSK 进行的所有内部和外部研究的行为准则,并概述了持续开发和提供有效和更安全药物的标准。克隆、基因改造和干细胞技术继续快速发展。现在,出于研究目的,对整个生物体(细菌、真菌、植物和动物)进行基因编辑已很常见。近年来,使用干细胞(见附件)治疗疾病和病症的情况也大幅增加。这些发展,加上从人类胚胎干细胞分离中产生的其他科学进步,已引起公众和监管机构对克隆、再生医学、干细胞研究和基因编辑方面的大量关注。本文概述了 GSK 对其中一些技术在医学研究中的重要性的看法,以及我们在适当情况下如何利用这些技术为尽可能多的人提供高质量且急需的医疗保健产品,以治疗和预防疾病。GSK 的看法是什么?

干细胞和可注射水凝胶
罗伊恩干细胞生物学和技术研究所,伊朗伊朗B冶金与材料工程系ACECR,ACECR,ACECR,伊朗工程学院,Semnan,Semnan,Semnan,伊朗大学,伊朗大学,伊朗c的遗传学系,伊斯兰教大学的材料,伊斯兰教大学,伊斯兰教大学,伊斯兰教材料,伊斯兰教材料,伊斯兰教材料马来西亚萨恩斯,工程校园,14300 Nibong tebal,槟城,马来西亚E E e化学工程系,东北大学,美国马萨诸塞州波士顿,美国马萨诸塞州波士顿,FioMaterials Innovations Innovations Innovations Innovations Innovations Innovations Innerovations Innervations研究中心,医学系,哈佛大学,美国马萨诸塞州波士顿,美国马萨诸塞州波士顿,美国,美国马萨诸塞州科技司,MASSACHSISTIUTTE为生物学启发的工程,哈佛大学,美国马萨诸塞州波士顿,美国科学与文化大学发展生物学系,ACECR,德黑兰,伊朗罗伊恩干细胞生物学和技术研究所,伊朗伊朗B冶金与材料工程系ACECR,ACECR,ACECR,伊朗工程学院,Semnan,Semnan,Semnan,伊朗大学,伊朗大学,伊朗c的遗传学系,伊斯兰教大学的材料,伊斯兰教大学,伊斯兰教大学,伊斯兰教材料,伊斯兰教材料,伊斯兰教材料马来西亚萨恩斯,工程校园,14300 Nibong tebal,槟城,马来西亚E E e化学工程系,东北大学,美国马萨诸塞州波士顿,美国马萨诸塞州波士顿,FioMaterials Innovations Innovations Innovations Innovations Innovations Innovations Innerovations Innervations研究中心,医学系,哈佛大学,美国马萨诸塞州波士顿,美国马萨诸塞州波士顿,美国,美国马萨诸塞州科技司,MASSACHSISTIUTTE为生物学启发的工程,哈佛大学,美国马萨诸塞州波士顿,美国科学与文化大学发展生物学系,ACECR,德黑兰,伊朗罗伊恩干细胞生物学和技术研究所,伊朗伊朗B冶金与材料工程系ACECR,ACECR,ACECR,伊朗工程学院,Semnan,Semnan,Semnan,伊朗大学,伊朗大学,伊朗c的遗传学系,伊斯兰教大学的材料,伊斯兰教大学,伊斯兰教大学,伊斯兰教材料,伊斯兰教材料,伊斯兰教材料马来西亚萨恩斯,工程校园,14300 Nibong tebal,槟城,马来西亚E E e化学工程系,东北大学,美国马萨诸塞州波士顿,美国马萨诸塞州波士顿,FioMaterials Innovations Innovations Innovations Innovations Innovations Innovations Innerovations Innervations研究中心,医学系,哈佛大学,美国马萨诸塞州波士顿,美国马萨诸塞州波士顿,美国,美国马萨诸塞州科技司,MASSACHSISTIUTTE为生物学启发的工程,哈佛大学,美国马萨诸塞州波士顿,美国科学与文化大学发展生物学系,ACECR,德黑兰,伊朗罗伊恩干细胞生物学和技术研究所,伊朗伊朗B冶金与材料工程系ACECR,ACECR,ACECR,伊朗工程学院,Semnan,Semnan,Semnan,伊朗大学,伊朗大学,伊朗c的遗传学系,伊斯兰教大学的材料,伊斯兰教大学,伊斯兰教大学,伊斯兰教材料,伊斯兰教材料,伊斯兰教材料马来西亚萨恩斯,工程校园,14300 Nibong tebal,槟城,马来西亚E E e化学工程系,东北大学,美国马萨诸塞州波士顿,美国马萨诸塞州波士顿,FioMaterials Innovations Innovations Innovations Innovations Innovations Innovations Innerovations Innervations研究中心,医学系,哈佛大学,美国马萨诸塞州波士顿,美国马萨诸塞州波士顿,美国,美国马萨诸塞州科技司,MASSACHSISTIUTTE为生物学启发的工程,哈佛大学,美国马萨诸塞州波士顿,美国科学与文化大学发展生物学系,ACECR,德黑兰,伊朗