XiaoMi-AI文件搜索系统
World File Search System极性相反

振动强耦合的共振理论增强了极性化学和光子模式寿命的作用
最近的实验表明,在振动强耦合(VSC)方面的极性子可以改变化学反应性。在这里,当将单个分子耦合到光腔时,我们介绍了VSC模化速率常数的完整理论,在该光腔中,人们了解了光子模式寿命的作用。分析表达表现出鲜明的共振行为,当腔频率与振动频率匹配时,达到最大速率常数。该理论解释了WHYVSC速率常数修饰与腔外振动的光谱非常相似。此外,我们讨论了VSC模化速率常数的温度依赖性。该分析理论与所有探索机制的运动层次(HEOM)模拟的数值确切层次方程(HEOM)非常吻合。最后,当考虑Fabry-Pérot腔内的平面动量时,我们讨论了正常发病率的共振条件。

rab6介导的突触囊泡前体的极化转运对于建立神经元极性和脑形成至关重要
神经元是由单个轴突和多个树突组成的高度极化细胞。轴突 - 树突极性对于正确的组织形成和脑功能至关重要。细胞内蛋白转运在神经元极性的建立中起重要作用。但是,极化运输的调节机制尚不清楚。在这里,我们表明Rab6是一种针对细胞内囊泡传统调节的小GTPase,在神经元极化和脑发育中起着关键作用。中枢神经系统特异性RAB6A/B双敲除(RAB6 DKO)两性的小鼠均表现出新皮质和小脑的严重发育不良。在Rab6 DKO新皮层中,神经元的轴突延伸受损会导致中间区发育不全。在体外,从性别中培养的神经元中Rab6a和Rab6b的缺失会导致与高尔基体相邻的突触囊泡前体(SVP)的异常积累,从而导致轴突延伸中的缺陷和Axon -Axon -dendrite Polarity的丧失。此外,Rab6 DKO会导致神经元中溶酶体的显着膨胀。总体而言,我们的结果表明,RAB6介导的SVP的极化转运对于神经元极化和随后的脑形成至关重要。

通过定向进化和计算建模构建富含 sp3 的多环化合物的生物催化策略
在 MD 轨迹中,分子重新定向,使得噻吩环位于血红素环 C 上方(1a 中为环 B),并且甲氧基的极性氧原子指向溶剂(图 6c)。相反,使用 6Me 取代的底物(2j)的 MD 模拟显示苯并噻吩环深深嵌入酶口袋中并远离溶剂(图 6c),其构象与底物 1a 和 5-OMe 底物不同。通过实验,酶对这些底物的活性遵循 2f(5-OMe)> 1a> 2j(6-Me)的顺序(图 3a)。由于该趋势与 MD 模拟确定的相应 TS 中苯并噻吩环对溶剂的暴露程度相关,我们假设底物

量子传感(Ⅰ):基础理论与方法
在物理和生命科学中具有广泛应用的固态量子传感器 ( 金刚石色心 -NV 氮原子空穴色心 ) ; 探索标准模型之外物理的量子传感器 ( 磁力仪和原子钟,囚禁的极性分子,自旋压缩,控制自旋退相 干,纠缠 ) ; 量子信息处理成为现实 ( 囚禁离子,约瑟夫森结 ) ; 增强型量子传感器的先进材料 ( 光晶格,固态量子缺陷,混合量子系统,拓扑材料 ) ; 用于暗区物理的量子传感器 ( 高 Q 值的射频或微波腔,基于超导干涉效应的高 Q 接收器 ) ; 基于原子干涉测量和光学原子钟的精密时空传感器 ( 量子纠缠 ( “压缩” ) 和量子控制 ( “动态解耦” )) 。
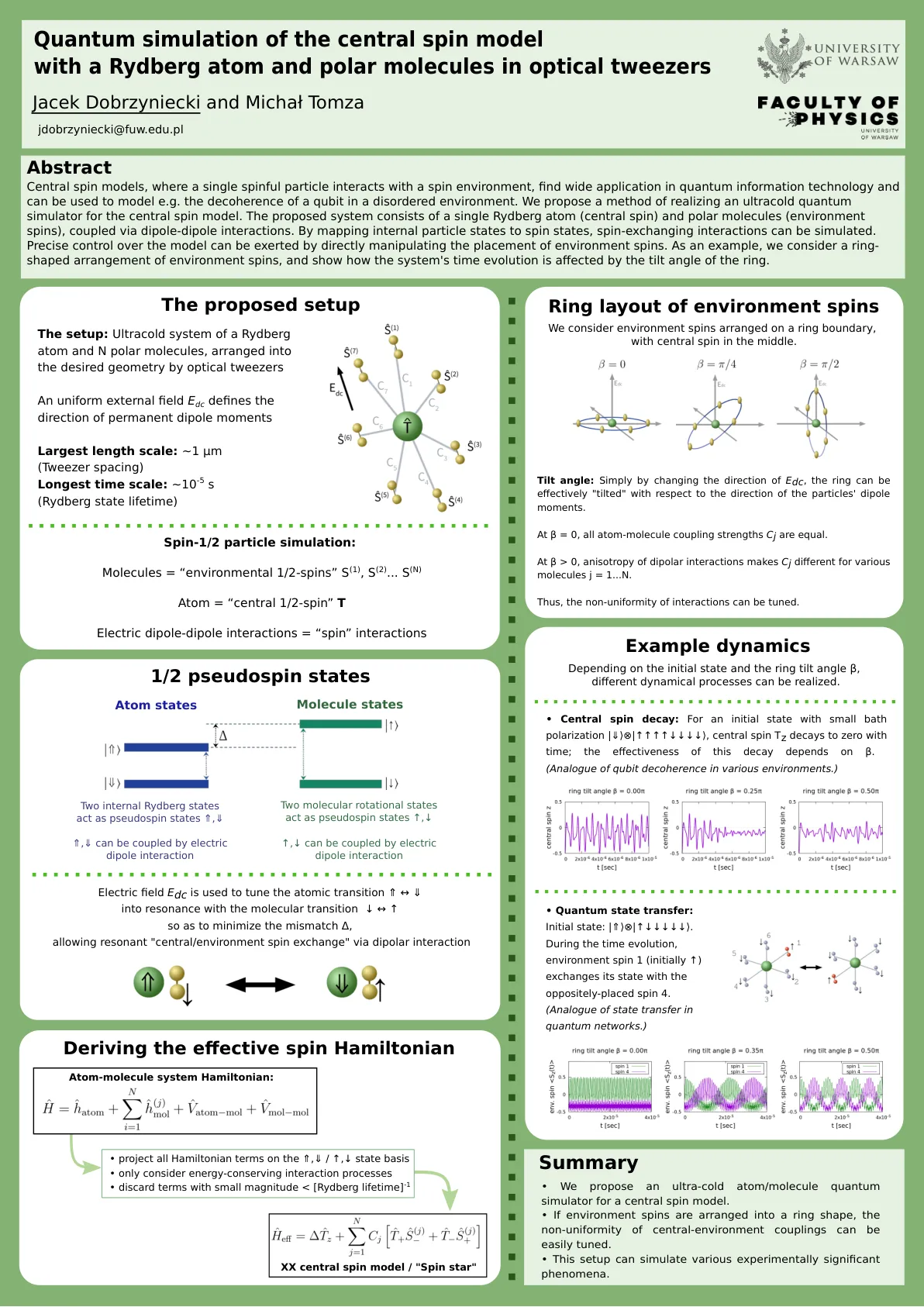
光镊中含有里德堡原子和极性分子的中心自旋模型的量子模拟
摘要 中心自旋模型(其中单个自旋粒子与自旋环境相互作用)在量子信息技术中得到广泛应用,并且可用于模拟无序环境中量子比特的退相干等。我们提出了一种实现中心自旋模型超冷量子模拟器的方法。所提出的系统由单个里德堡原子(中心自旋)和极性分子(环境自旋)组成,它们通过偶极-偶极相互作用耦合。通过将内部粒子状态映射到自旋状态,可以模拟自旋交换相互作用。可以通过直接操纵环境自旋的位置来精确控制模型。作为示例,我们考虑环境自旋的环形排列,并展示系统的时间演化如何受到环的倾斜角的影响。

脑脊液神经胶质原纤维酸性蛋白与淀粉样蛋白蛋白相反,与亨廷顿氏病
a Department of Medicine, Neurology, V ¨ asterås Central Hospital, V ¨ asterås, Sweden b Department of Medical Sciences, Neurosurgery, Uppsala University, Uppsala, Sweden c Department of Psychiatry and Neurochemistry, Institute of Neuroscience and Physiology, the Sahlgrenska Academy at the University of Gothenburg, M ¨ olndal,瑞典Sahlgrenska大学医院,瑞典E Olndal,瑞典E部,UCL神经退行性疾病系,UCL神经病学研究所,英国伦敦皇后广场,英国伦敦皇后广场,UCL伦敦UCL,UCL,UCL,UCL,UCL,UCL,UCL,UCL,UCL,UCL研究所,英国G香港神经变性疾病,哥伦比亚省神经退行性疾病中心,神经科学,Sahlgrenska学院,哥德堡大学,哥德堡大学,瑞典I临床神经科学系,Karolinska Institutet,斯德哥尔摩,斯德哥尔摩,瑞典J医学科学系

超级电容器及其革命性能源存储和电力输送的潜力
传统电容器是双端无源电气元件,以电场的形式静电存储能量。它们由两个导电表面(也称为电极)组成,由电介质或绝缘体隔开。当在电容器上施加电压时,电子会向其中一个极板迁移,在其上产生净正电荷,并排斥另一个极板上的电子。由于相反电荷之间的静电吸引力,正电荷和负电荷保留在极板上。极板之间的绝缘体可防止因电位差而导致的任何电荷迁移,因此没有电流流过电容器。这在两个极板之间产生了电场,该电场一直持续到外部端子带电、短路或施加在电容器上的电压极性发生变化为止。这一特性是电容器储能能力的本质,即使电容器与电压源断开连接,电压仍会保持。

可持续可伸缩电池用于下一代可穿戴设备
在大规模模拟中,由于量子方法的数值成本很高,原子之间的相互作用通常不能从第一个原理计算。相反,它们通常是使用力ELDS(FFS)对势能的物理动机功能形式进行建模的,并进行参数化以匹配从头算能的能量和/或再现实验数据。最广泛的FF是所谓的经典力eLS(例如Amber 1或Charmm 2),它们结合了XED-Charge Colomb电位和Lennard-Jones的相互作用来模拟分子间电位。这些模型在数值上非常有效,可以在长期尺度上模拟非常大的系统。然而,它们的简单功能形式缺乏极化和多体效应,这对于正确描述某些系统至关重要(例如在极性溶剂,PI堆叠或复杂的蛋白质结构中溶剂化3)。更先进的力量eelds - 例如Amoeba,4 TTM,5

CUC1/生长素基因模块将十字花科植物的细胞极性与组织生长模式和叶片形状多样性联系起来
空间分布的基因活动如何转化为细胞极性和生长模式,从而产生多种形式的多细胞真核生物,这一点仍不清楚。在这里,我们表明,转录因子杯形子叶 1 (CUC1) 的物种特异性表达是两种相关植物物种之间叶形差异的关键决定因素。通过结合延时成像、遗传学和建模,我们发现 CUC1 充当极性开关。该开关通过转录激活影响生长素转运蛋白极性的激酶来调节叶形,生长素转运蛋白通过与激素生长素的反馈来模式化叶片生长。因此,我们发现了一种机制,通过将物种特异性转录因子表达与细胞水平极性和生长联系起来,跨越生物尺度,形成不同的叶形。

A和Tau蛋白在阿尔茨海默氏病中的作用
抽象实现具有窄带发射和高颜色纯度的高发光有机发光设备(OLEDS)在各种光电领域都很重要。激光显示由于其最终的视觉体验而在下一代展示技术中表现出了出色的优势,但这仍然是一个巨大的挑战。在这里,我们开发了一种新型的基于OLED的有机单晶。通过将有机激子状态与光学微腔内强烈耦合,我们从极性的OLED(OPLEDS)中获得了Polariton电致工(EL)发射,具有较高的亮度,窄带发射,高色纯度,高极性,高极性以及出色的光学泵送极性元素Laser。此外,我们通过理论分析评估了电泵浦极性激光的潜力,并提供了可能的解决方案。这项工作提供了一种强大的策略,具有材料 - 设备组合,为电动有机单晶的极性发光设备和可能的激光器铺平了道路。