XiaoMi-AI文件搜索系统
World File Search System氨基糖

胰岛素调节的氨基肽酶的水平降低(...
背景:胰岛素调节的氨基肽酶(IRAP)参与胰岛素敏感性和葡萄糖代谢,在2型糖尿病的病理生理中很重要。血清IRAP水平与2型糖尿病和胰岛素抵抗密切相关。这项研究的目的是评估IRAP水平作为妊娠糖尿病(GDM)妇女早期诊断和管理胰岛素抵抗的潜在生物标志物。方法:这项队列研究包括40名GDM女性和40名健康妊娠女性。母体血清IRAP水平,并在两组之间进行比较。结果:与对照组(0.92±0.10 ng/ml)相比,GDM组的平均血清IRAP水平明显降低(0.73±0.12 ng/ml)(p = 0.001)。成对比较表明,经过修饰和胰岛素治疗的GDM亚组的血清IRAP水平明显低于对照组(分别为p <0.017和p <0.017)。血清IRAP水平与禁食葡萄糖,胰岛素,稳态模型耐药性(HOMA-IR)水平和血红蛋白A1C(HBA1C)(r = –0.541,P = 0.001; r = 0.001; r = –0.447,p = 0.001; r = 0.584,p = –0.584,P = –0.584,P = 0.001; R = 0.001; 0.001)。最佳血清IRAP截止值计算为0.857 ng/ml,灵敏度为85%,对于GDM的预测,特异性为80%(p = 0.001)。结论:被诊断为GDM的孕妇的血清IRAP水平明显低于健康孕妇。此外,血清IRAP水平与胰岛素,HBA1C和HOMA-IR的水平负相关。这些发现表明,低血清IRAP水平可能是预测GDM的新型生物标志物。临床试验注册:该研究已在https://classic.clinicaltrials.gov/上注册(注册号:NCT06716320)。

疟疾氨基肽酶评论-修订版.pdf
重复使用 存放在 White Rose Research Online 中的项目受版权保护,保留所有权利,除非另有说明。您可以下载和/或打印这些项目用于个人学习,或国家版权法允许的其他行为。出版商或其他权利持有人可能允许进一步复制和重复使用全文版本。这由 White Rose Research Online 记录中该项目的许可信息指示。
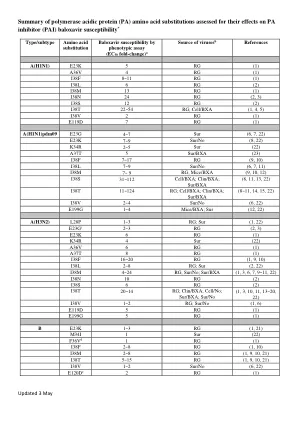
摘要 - 聚合酶 - 酸性 - (PA) - 蛋白质 - 氨基 -
* PA中的其他氨基酸取代,在参考文献1(Omoto S等,2018)和#2(Hashimoto T等,2020年)中研究了Baloxavir易感性没有变化的其他氨基酸取代。通过基于细胞培养的测定法评估(焦点,斑块或屈服分析,高含量成像中和(提示)和ViroDot分析)。EC 50倍变化。b细胞,细胞培养;临床试验;小鼠,鼠标模型; RG,反向遗传学; SUR,监视研究; BXA,在Baloxavir压力下选出的取代;不,Baloxavir不使用。c e23g(T0831)。通过表型测定测试了带有E23G的RG病毒。d对应于A36V A型A型PA中的A36V。 E对应于A型A型PA中的E119D。参考文献1。Omoto S,Speranzini V,Hashimoto T,Noshi T,Yamaguchi H,Kawai M,Kawaguchi K,Uehara T,Shishido T,Naito A,Naito A,Cusack S.2018。通过核酸内切酶抑制剂Baloxavir maroxil诱导的流感病毒变体的表征。SCI REP 8:9633。2。Hashimoto T,Baba K,Inoue K,Okane M,Hata S,Shishido T,Naito A,Wildum S,Omoto S.2020。在Baloxavir Marboxil的临床试验中检测到的流感病毒的三聚体RNA聚合酶复合物中氨基酸取代的全面评估。流感其他呼吸病毒DOI:10.1111/irv.12821。3。ince WL,Smith FB,O'Rear JJ,Thomson M.2020。J Infect DIS 222:957-961。 4。 2018。J Infect DIS 222:957-961。4。2018。治疗 - 伴随流感病毒聚合酶酸性取代率与Balosavir Maroxavir Marboxil试验中的i38中的i38中的酸性取代相关。Noshi T, Kitano M, Taniguchi K, Yamamoto A, Omoto S, Baba K, Hashimoto T, Ishida K, Kushima Y, Hattori K, Kawai M, Yoshida R, Kobayashi M, Yoshinaga T, Sato A, Okamatsu M, Sakoda Y, Kida H, Shishido T, Naito A.Baloxavir酸的体外表征,Baloxavir酸是一种流感病毒聚合酶PA亚基的第一类帽依赖性内切酶抑制剂。抗病毒Res 160:109-117。5。Takashita E,Morita H,Ogawa R,Nakamura K,Fujisaki S,Shirakura M,Kuwahara T,Kishida N,Watanabe S,Odagiri T.2018。流感病毒对新型帽依赖性核酸内切酶抑制剂baloxavir maroxil的敏感性。前微生物9:3026。6。Gubareva LV,Mishin VP,Patel MC,Chesnokov A,Nguyen HT,De La Cruz J,Spencer S,Spencer S,Campbell AP,Sinner M,Reid H,Reid H,Garten R,Katz JM,Katz JM,Fry AM,Barnes J,Barnes J,Wentworth DE。 2019。 评估在2016/17和2017/18季节在美国循环的流感病毒的Baloxavir敏感性。 欧元监视24:1800666。 7。 Takashita E, Daniels RS, Fujisaki S, Gregory V, Gubareva LV, Huang W, Hurt AC, Lackenby A, Nguyen HT, Pereyaslov D, Roe M, Samaan M, Subbarao K, Tse H, Wang D, Yen HL, Zhang W, Meijer A. 2020。 全球关于人流感病毒对神经氨酸酶抑制剂和cap依赖性核酸内切酶抑制剂Baloxavir的敏感性的更新,2017- 2018年。 抗病毒Res 175:104718。 8。 2020。Gubareva LV,Mishin VP,Patel MC,Chesnokov A,Nguyen HT,De La Cruz J,Spencer S,Spencer S,Campbell AP,Sinner M,Reid H,Reid H,Garten R,Katz JM,Katz JM,Fry AM,Barnes J,Barnes J,Wentworth DE。2019。评估在2016/17和2017/18季节在美国循环的流感病毒的Baloxavir敏感性。欧元监视24:1800666。7。Takashita E, Daniels RS, Fujisaki S, Gregory V, Gubareva LV, Huang W, Hurt AC, Lackenby A, Nguyen HT, Pereyaslov D, Roe M, Samaan M, Subbarao K, Tse H, Wang D, Yen HL, Zhang W, Meijer A.2020。全球关于人流感病毒对神经氨酸酶抑制剂和cap依赖性核酸内切酶抑制剂Baloxavir的敏感性的更新,2017- 2018年。抗病毒Res 175:104718。8。2020。Takashita E, Abe T, Morita H, Nagata S, Fujisaki S, Miura H, Shirakura M, Kishida N, Nakamura K, Kuwahara T, Mitamura K, Ichikawa M, Yamazaki M, Watanabe S, Hasegawa H, Influenza Virus Surveillance Group of J.流感A(H1N1)PDM09病毒,由于未经Baloxavir治疗的儿童检测到PA E23K替代而表现出对Baloxavir的敏感性降低。抗病毒Res 180:104828。 9。 Koszalka P,Tilmanis D,Roe M,Vijaykrishna D,Hurt AC。 2019。 亚太地区流感病毒的Baloxavir Marboxil易感性,2012- 2018年。 抗病毒Res 164:91-96。 10。 Jones JC,Pascua PNQ,Fabrizio TP,Marathe BM,Seiler P,Barman S,Webby RJ,Webster RG,Govorkova EA。 2020。 流感和B病毒具有降低的Baloxavir敏感性显示器的体外适应性减弱,但保留了雪貂的可传播性。 Proc Natl Acad Sci U S A 117:8593-8601。 11。 Chesnokov A,Patel MC,Mishin VP,De La Cruz JA,Lollis L,Nguyen HT,Dugan V,Wentworth DE,Gubareva LV。 2020。 季节性流感A病毒的复制适应性,对Baloxavir的敏感性降低。 J Infect DIS 221:367-371。 12。 Kiso M,Yamayoshi S,Murakami J,Kawaoka Y. 2020。 Baloxavir Marboxil治疗感染了流感病毒的裸小鼠。 J Infect Dis 221:1699-1702。 13。 Sato M,Takashita E,Katayose M,Nemoto K,Sakai N,Hashimoto K,HosoyaM.2020。 J Infect DIS 222:121-125。 14。 J Infect DIS 221:63-70。 15。 2020。 16。抗病毒Res 180:104828。9。Koszalka P,Tilmanis D,Roe M,Vijaykrishna D,Hurt AC。2019。亚太地区流感病毒的Baloxavir Marboxil易感性,2012- 2018年。抗病毒Res 164:91-96。 10。 Jones JC,Pascua PNQ,Fabrizio TP,Marathe BM,Seiler P,Barman S,Webby RJ,Webster RG,Govorkova EA。 2020。 流感和B病毒具有降低的Baloxavir敏感性显示器的体外适应性减弱,但保留了雪貂的可传播性。 Proc Natl Acad Sci U S A 117:8593-8601。 11。 Chesnokov A,Patel MC,Mishin VP,De La Cruz JA,Lollis L,Nguyen HT,Dugan V,Wentworth DE,Gubareva LV。 2020。 季节性流感A病毒的复制适应性,对Baloxavir的敏感性降低。 J Infect DIS 221:367-371。 12。 Kiso M,Yamayoshi S,Murakami J,Kawaoka Y. 2020。 Baloxavir Marboxil治疗感染了流感病毒的裸小鼠。 J Infect Dis 221:1699-1702。 13。 Sato M,Takashita E,Katayose M,Nemoto K,Sakai N,Hashimoto K,HosoyaM.2020。 J Infect DIS 222:121-125。 14。 J Infect DIS 221:63-70。 15。 2020。 16。抗病毒Res 164:91-96。10。Jones JC,Pascua PNQ,Fabrizio TP,Marathe BM,Seiler P,Barman S,Webby RJ,Webster RG,Govorkova EA。2020。流感和B病毒具有降低的Baloxavir敏感性显示器的体外适应性减弱,但保留了雪貂的可传播性。Proc Natl Acad Sci U S A 117:8593-8601。11。Chesnokov A,Patel MC,Mishin VP,De La Cruz JA,Lollis L,Nguyen HT,Dugan V,Wentworth DE,Gubareva LV。2020。季节性流感A病毒的复制适应性,对Baloxavir的敏感性降低。J Infect DIS 221:367-371。 12。 Kiso M,Yamayoshi S,Murakami J,Kawaoka Y. 2020。 Baloxavir Marboxil治疗感染了流感病毒的裸小鼠。 J Infect Dis 221:1699-1702。 13。 Sato M,Takashita E,Katayose M,Nemoto K,Sakai N,Hashimoto K,HosoyaM.2020。 J Infect DIS 222:121-125。 14。 J Infect DIS 221:63-70。 15。 2020。 16。J Infect DIS 221:367-371。12。Kiso M,Yamayoshi S,Murakami J,Kawaoka Y. 2020。 Baloxavir Marboxil治疗感染了流感病毒的裸小鼠。 J Infect Dis 221:1699-1702。 13。 Sato M,Takashita E,Katayose M,Nemoto K,Sakai N,Hashimoto K,HosoyaM.2020。 J Infect DIS 222:121-125。 14。 J Infect DIS 221:63-70。 15。 2020。 16。Kiso M,Yamayoshi S,Murakami J,Kawaoka Y.2020。Baloxavir Marboxil治疗感染了流感病毒的裸小鼠。 J Infect Dis 221:1699-1702。 13。 Sato M,Takashita E,Katayose M,Nemoto K,Sakai N,Hashimoto K,HosoyaM.2020。 J Infect DIS 222:121-125。 14。 J Infect DIS 221:63-70。 15。 2020。 16。Baloxavir Marboxil治疗感染了流感病毒的裸小鼠。J Infect Dis 221:1699-1702。13。Sato M,Takashita E,Katayose M,Nemoto K,Sakai N,Hashimoto K,HosoyaM.2020。 J Infect DIS 222:121-125。 14。 J Infect DIS 221:63-70。 15。 2020。 16。Sato M,Takashita E,Katayose M,Nemoto K,Sakai N,Hashimoto K,HosoyaM.2020。J Infect DIS 222:121-125。14。J Infect DIS 221:63-70。15。2020。16。在2018-2019流感季节治疗流感A的儿童后,检测Baloxavir Marboxil易感性降低的变体。Checkmahomed L,M'Hamdi Z,Carbonneau J,Venable MC,Baz M,Abed Y,Boivin G.2020。抗性抗性聚合酶酸I38T取代对当代流感A(H1N1)PDM09和A(H3N2)菌株的适应性的影响。Imai M, Yamashita M, Sakai-Tagawa Y, Iwatsuki-Horimoto K, Kiso M, Murakami J, Yasuhara A, Takada K, Ito M, Nakajima N, Takahashi K, Lopes TJS, Dutta J, Khan Z, Kriti D, van Bakel H, Tokita A, Hagiwara H, Izumida N,Kuroki H,Nishino T,Wada N,Koga M,Adachi E,Jubishi D,木谷H,Kawaoka Y.流感A的变体降低了对日本患者分离的Baloxavir敏感性的变体,并通过呼吸道液滴进行拟合。NAT微生物5:27-33。 Takashita E, Kawakami C, Morita H, Ogawa R, Fujisaki S, Shirakura M, Miura H, Nakamura K, Kishida N, Kuwahara T, Mitamura K, Abe T, Ichikawa M, Yamazaki M, Watanabe S, Odagiri T, On Behalf Of The Influenza VirusNAT微生物5:27-33。Takashita E, Kawakami C, Morita H, Ogawa R, Fujisaki S, Shirakura M, Miura H, Nakamura K, Kishida N, Kuwahara T, Mitamura K, Abe T, Ichikawa M, Yamazaki M, Watanabe S, Odagiri T, On Behalf Of The Influenza Virus

脂肪糖诱导的糖尿病大鼠脂肪糖诱导的根尖牙周炎对根尖的炎症
抽象目的是通过白介素6(IL-6)(IL-6)和肿瘤坏死因子α(TNF-A)的免疫组织化学分析来评估根本炎症的抽象目标。大鼠上糖尿病模型的材料和方法是由链蛋白酶(STZ)诱导的。将15只大鼠注入低剂量STZ 5天,等待5天,直到血糖水平稳定,并通过数字糖仪测量了300 mg/dL以上的测量。LPS用于诱导根尖牙周炎。执行访问腔后,在麻醉下,在大鼠的第一摩尔的根管空间上进行了牙髓和根管的灭绝。LPS在牙髓和根管空间中诱导了1 mg/ml剂量。顶端牙周炎预计是14天后的,然后将大鼠随机分配给三组。第一个组在诱导后14天终止并用作对照。诱导后28天观察到第二组,并且在诱导后42天观察到第三组。IL-6和TNF- A表达。使用单向方差分析分析统计分析数据,并继续进行HOC Tukey HSD测试。明显的能力。结果LPS在对照组(14天),28天和42天观察的糖尿病大鼠中诱导顶端牙周炎,显示IL-6和TNF-A的表达显着增加。对照组和观察到的组之间存在显着差异(p <0.05)。在14和28天(p> 0.05)时,IL-6在顶端区域的表达并不显着,但在42天时显着增加(p <0.05)。14天后,TNF-A在顶端区域中的表达显着增加(p <0.05),并在28和42天保持稳定(p> 0.05)。

针对己糖激酶 2 进行口腔癌治疗
针对己糖激酶 2 (HK2) 的小分子抑制剂的研发极大地吸引了癌症药物研发领域的注意力。然而,针对己糖激酶 (HK) 特定异构体的选择性抑制剂的研发仍然是一项艰巨的挑战。在这里,我们提出了一种多药效团建模方法,用于设计针对 HK2 的配体,对 FaDu 和 Cal27 口腔癌细胞系具有显着的抗增殖作用。分子动力学 (MD) 模拟表明,原型配体对 HK2 表现出更高的亲和力。除此之外,我们提出了一种可持续的合成途径:一种环保的单步工艺,通过在无过渡金属条件下以优异的产率在环境温度下将酯与胺直接酰胺化来促进,然后采用避免柱层析分离技术分离已鉴定的先导生物活性化合物(H2),该化合物表现出细胞周期停滞和细胞凋亡。我们观察到 HK2 的抑制导致线粒体膜电位丧失和线粒体自噬增加,这是一种潜在的抗癌作用机制。先导 H2 还减少了球体的生长。总之,这些结果表明,原型先导化合物具有抗癌潜力的 HK2 抑制的概念验证。


与糖相关的知识、态度和实践......
本执行摘要是 2023 年阿曼苏丹国营养相关知识、态度和实践调查的一部分。该调查由卫生部营养司在世界卫生组织和 Al Jisr 基金会的支持下进行。特别感谢参与本次调查的青少年和成年人,以及从阿曼苏丹国所有省份收集数据的实地工作人员。本执行摘要由 Salima Almamary 博士、Halima Alghannami 博士、Saleh Al Shammakhi 先生、Ibtisam Al Ghammari 女士、Nicolai Petry 博士和 James P Wirth 博士编写。

用于顺势疗法的 Mulk 糖
除非另有规定,试验中应使用纯化学品,当打算使用水作为试剂时,应使用蒸馏水(见 IS 1070)。“纯化学品”指不含影响测试结果的杂质的化学品。 4 保质期 建议保质期为 2 年,以内部稳定性研究的数据为准。 5 取样 应按照 IS 1000 的规定抽取产品的代表性样品并测试其是否符合本标准。 6 包装和标志 6.1 包装 6.1.1 零售包装 产品应装在干净、完好的金属容器(见 IS 11078)或玻璃容器或惰性塑料容器,如 PET 和 HDPE 罐(见 IS 19084)或食品级软包装中,该包装由薄膜或纸板、纸、聚乙烯、聚酯金属化薄膜或铝箔等任何基材的组合制成,以防止其变质。如果使用塑料材料,只能使用食品级塑料(见 IS 10171)。6.1.2 散装包装产品可以 25 公斤的量装入食品级聚乙烯袋中(见 IS 10171),厚度至少为 0.05 毫米。袋子应该通过缝纫或热封进行适当封闭。

患病率和风险新的糖尿病 - 糖 -
COVID-19的大流行今天仍然是全球紧急状态,经过将近三年的传播,它的传播已经远远无法克服。SARS-COV-2感染后有几种临床表现,包括中期和长期Covid-19,它们主要需要特别注意,主要是因为它们与其他合并症的关系(1)。已经观察到,糖尿病,高血压和肥胖等合并症的高龄和存在与Covid-19的最严重形式有关(2)。由于SARS-COV-2感染后的第一个有关Covid-19的报告的报告是关于COVID-19的流行病糖尿病患者的急性并发症风险的增加,甚至有高风险,即使随后发展出所谓的“长期共vid综合征”的各种症状,也有多种症状(3)(3)。越来越多的证据还表明,长期covid的众多临床异常甚至可能扩展到新的发作糖尿病(4)。因此,COVID-19与糖尿病之间的关系可以被视为双向。一方面,糖尿病患者感染SARS-COV-2的风险增加,而血糖控制不足和慢性血管并发症患者的风险尤其尤其高,可能会出现最严重的疾病形式(5)。另一方面,考虑到COVID-19感染的患者,考虑到与糖尿病相关的急性并发症的风险也增加了(6)。已经证明,与ACE-2受体结合后,SARS-COV-2病毒在人类细胞中进入(9)。在这种关系基础的不同机制中,血管紧张素转换酶2(ACE-2)受体可能在关键的代谢器官和包括胰腺β细胞在内的关键代谢器官和组织中的广泛定位,可能具有作用(7,8)。越来越多的证据表明,感染SARS-COV-2的人患有糖尿病的风险增加,并且在疾病的急性阶段中使用抗血糖疗法的事件(4,10),根据糖尿病本身的常规危险因素的风险增加,风险增加了风险。然而,该证据并不普遍,尤其是当SARS-COV-2感染后糖尿病发作时(11)。根据此背景,我们的系统审查和荟萃分析的目的是:(1)评估诊断至少60天后感染SARS-COV-2的患者中糖尿病患病率和任何类型的葡萄糖代谢异常。 (2)至少在SARS-COV-2感染后至少60天后估计新发作糖尿病的比例,以排除继发于类固醇继发或与疾病急性期使用或相关的所有高血糖。

标题:减少能源,钠,糖,...
2013 2014 2015 2016 2016 2017 2019 2019家庭尺寸4.2 4.2 4.2 4.2 4.2 4.2 4.3 4.4 4.5(1.6)(1.6)(1.6)(1.6)(1.7)(1.7)(1.7)(1.7)(1.7)主要购物者的年龄47.9 48.9 48.3 48.2 48.6 48.6 48.6 49.3 49.3 49.4 49.4 49.4 49.4 49.4 49.4 49.4 49.4 49.4 49.4 49.4(14.5)(14.5)(14.7)(14.7)(14.7)(14.7) (15.5) (15.5) Number of cars 0.5 0.6 0.7 0.7 0.7 0.7 0.7 (0.7) (0.7) (0.8) (0.7) (0.7) (0.7) (0.8) Number bathrooms 1.3 1.3 1.3 1.3 1.3 1.3 1.3 (0.6) (0.6) (0.6) (0.6) (0.6) (0.6) (0.7) Number bedrooms 2.9 3.0 3.1 3.0 3.0 3.0 3.1 (1.0) (2.4) (1.5) (1.0) (1.0) (1.0) (1.0) Household head's education Lower than high school 42% 40% 37% 32% 31% 28% 27% High school 37% 38% 40% 42% 42% 45% 44% Higher than high school 21% 22% 23% 25% 27% 28% 29% Socioeconomic status (SES) ABC1 (highest) 8% 8% 8% 8% 8% 8% 7% C2 17% 17% 17% 16% 16% 17% 16% C3 27% 27% 27% 27% 27% 27% 28% DE (lowest) 49% 49% 49% 49% 49% 48% 49% Region North 13% 13% 12% 13% 13% 13% 13% Center 64% 63% 63% 63% 63% 62% 62% South 24% 24% 24% 24% 24% 25% 25%的家庭月数量。11,636 23,455 23,292 23,385 22,762 22,684 11,177注:样本将于2013年7月开始,于2019年6月结束。SES的定义基于智利的关联229