XiaoMi-AI文件搜索系统
World File Search System活性

CpG DNA的免疫活性及应用前景
[20]MccluskIe MJ,Davis H1. . cpG DNA㈣potent enhancer 0f 8ystemi㈨d洲osal㈣une respo—s ngainst hepatitIs B surfa㈣ntIg洲“h mtrana蛆l admini8tration to mice J I响u—

肿瘤免疫检查点PD-1/PD-L1抑制剂的中药活性成分研究进展
表没食子茶素没食子酸酯 (EGCG) ,是茶多酚 中最有效的活性成分,属于儿茶素类化合物。 EGCG 具有抗菌、抗炎、抗病毒、抗肿瘤等作用 [ 21 ] 。有研 究报道,在非小细胞肺癌中, EGCG 能够抑制 IFN- γ 和表皮生长因子( EGF )诱导的 PD-L1 的表达。 EGCG 和绿茶提取物能够抑制 A549 人肺癌细胞 中 JAK2/STAT1 信号通路,从而减少由 IFN-γ 诱导 的 PD-L1 mRNA 以及蛋白质的表达水平;同时通 过抑制 EGF 受体 /AKT 信号通路,使 EGF 诱导的 PD-L1 的表达降低。在腹腔注射 4- 甲基亚硝胺基 - 1- ( 3- 吡啶基) -1- 丁酮 (NNK) 诱导的小鼠肺癌模型 中,小鼠的饮用水中加入 0.3% 的绿茶提取物,可以 降低每只小鼠的平均肿瘤数目和 70% PD-L1 的阳 性细胞率。在 F10-OVA 黑色瘤细胞和肿瘤特异 性 CD3+T 细胞共培养模型中, EGCG 能够使 F10- OVA 细胞的 PD-L1 mRNA 的表达降低,并且可以 恢复肿瘤特异性 CD3+T 细胞 IL-2 mRNA 的表 达 [ 22 ] 。这些结果表明, EGCG 是 PD-L1 的有效抑制 剂,具有抑制 EGFR/Akt 和 IFNR/JAK2/STAT1 通 路的潜力。
活性A
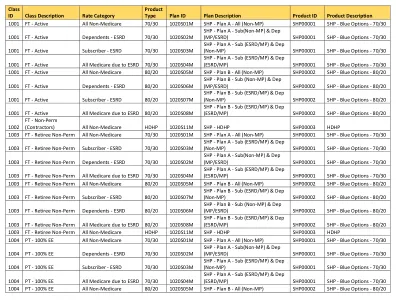
SHP - 计划 B - 附属 (ESRD/MP) 和支助 (ESRD/MP) SHP00002 SHP - 蓝色选项 - 80/20 1014 COBRA COBRA - 计划 A 70/30 1020S09M SHP - 计划 A 直接支付 SHP00001 SHP - 蓝色选项 - 70/30 1014 COBRA COBRA - 计划 B 80/20 1020S10M SHP - 计划 B 直接支付 SHP00002 SHP - 蓝色选项 - 80/20 1014 COBRA COBRA - HDHP HDHP 1020S11M SHP - HDHP SHP00003 HDHP



量化活性胶体的非平衡活性
活性胶体是能够自推进的粒子,能在微观尺度上将化学能转化为定向的机械运动 [1]。它们已成为活性物质领域的典范,因为它们表现出相变 [3] 和动态结晶 [4] 等突发行为 [2],也是研究非平衡微观热机的基础 [5–8]。人们已投入大量精力开发一个框架来理解活性物质,并将其与随机热力学联系起来 [9–13],将经典热力学的概念扩展到非平衡系统和个体轨迹。这种方法的一个普遍局限性是,由于热噪声和活性噪声不能沿轨迹明确分离,因此熵的产生不能完全推断 [14]。尽管如此,随机热力学有潜力推动该领域从研究活性物质的特定现象学模型转向开发驱动活性系统的通用热力学框架。活性物质系统在广泛的空间和时间尺度上无处不在[15–17]。在纳米尺度上,单个分子可以充当活性物质[18, 19];在研究最深入的微观尺度上,生物和合成系统起着活性物质的作用[20–24];在中尺度和更大尺度上,动物[25]、机器人[26]、人类群体[27]等作为活性物质运行。所有这些系统所受控的底层物理过程千差万别,如湿与干[16, 28]、欠阻尼与过阻尼[29–32]、热与非热[33–35]等。然而,它们都有一个重要的共同点——非平衡动力学的出现是因为活性物质系统中的每个元素都会消耗能量并耗散


开发一种测量肿瘤组织和血液中 DNA 损伤修复活性的方法
对于修复DNA双链断裂的同源重组修复(注4)异常的肿瘤,PARP抑制剂和铂类抗癌药物被认为有效。此外,同源重组修复所需分子的基因异常会导致遗传性乳腺癌和卵巢癌综合征。 东北大学发育、衰老和癌症研究所肿瘤生物学系的助理教授 Yoshino Yuki 和教授 Chiba Natsuko 此前开发了一种测量癌细胞中同源重组修复能力的方法,并证明了其准确性(Sci Rep 2019、Cancer Res Commun 2021)。 现在,由研究生 Tokikazu Motonari 和东京大学医学院乳腺和内分泌外科系教授 Takanori Ishida 组成的合作研究小组成功开发出一种测量小鼠肿瘤组织和血液来源的淋巴母细胞同源重组修复活性的方法。 有人提出,应用这些方法可能可以预测癌症治疗药物的有效性以及诊断遗传性乳腺癌和卵巢癌综合征。


肿疡溶解性ウイルスによる抗肿疡免疫の活性化と...
15 -ürCatamama Y.等,J. Immunol。,200,2987-2999 2018-16-16-ürT.等,摩尔。 ther。 Oncolytics,12,162-172 2018-17-17-17-17-17-17-1等,等,Nat。 Society。,9,251 2018 18 -Low E.等,当前。 opine。 Biochnol。,65,25-36 (2020-19 – 19 – Mahalingam D.等,Clin。 res。,26,71-81 2020-20-Sun L.等,J。Imunother。 癌症,6,6,36 USBAND 21-储存C.和Al。,Clin。 res。,25,290-299!hang Y.和al。,mol。 ther。,27,1906-1 ther。 Oncolytics,26,265-274 2022-64-Emprot K.和Al。,Hum。 基因。15 -ürCatamama Y.等,J.Immunol。,200,2987-2999 2018-16-16-ürT.等,摩尔。 ther。 Oncolytics,12,162-172 2018-17-17-17-17-17-17-1等,等,Nat。 Society。,9,251 2018 18 -Low E.等,当前。 opine。 Biochnol。,65,25-36 (2020-19 – 19 – Mahalingam D.等,Clin。 res。,26,71-81 2020-20-Sun L.等,J。Imunother。 癌症,6,6,36 USBAND 21-储存C.和Al。,Clin。 res。,25,290-299!hang Y.和al。,mol。 ther。,27,1906-1 ther。 Oncolytics,26,265-274 2022-64-Emprot K.和Al。,Hum。 基因。Immunol。,200,2987-2999 2018-16-16-ürT.等,摩尔。ther。Oncolytics,12,162-172 2018-17-17-17-17-17-17-1等,等,Nat。Society。,9,251 2018 18 -Low E.等,当前。opine。Biochnol。,65,25-36 (2020-19 – 19 – Mahalingam D.等,Clin。res。,26,71-81 2020-20-Sun L.等,J。Imunother。癌症,6,6,36 USBAND 21-储存C.和Al。,Clin。res。,25,290-299!hang Y.和al。,mol。ther。,27,1906-1ther。Oncolytics,26,265-274 2022-64-Emprot K.和Al。,Hum。基因。