XiaoMi-AI文件搜索系统
World File Search System突可

单纤维激光器中的孤子运动
摘要:很少有模型可以研究人类中枢神经系统中的神经突损伤。我们在这里使用多巴胺能LUHMES神经元来建立一个培养系统,该系统允许(i)观察高度富集的神经突,(ii)为生化研究制备神经突级分的生化研究,以及(iii)轴突造口后神经酸盐标记物和代谢物的测量。luhmes的球体,在培养皿中镀以数千m的长度,而所有somata均保持聚集。这些培养物可以轻松地观察活的神经突或固定神经突。纯神经突(NOC)。通过确定其蛋白质和RNA含量来说明这种培养物的潜在应用。例如,线粒体TOM20蛋白高度丰富,而核组蛋白H3则没有。同样,在相对较高的水平上发现了线粒体编码的RNA,而在NOC中,组蛋白或神经元核标记NEUN(RBFOX3)的mRNA相对耗尽。NOC的另一种潜在用途是神经突变性的研究。为此,开发了一种量化神经突完整性的算法。使用此工具,我们发现烟酰胺的添加大大降低了神经突变性。另外,NOC中Ca 2+的螯合延迟了变性,而Calpains的抑制剂也没有作用。因此,NOC被证明适用于生化分析和在定义的切割损伤后研究变性过程。

人H-铁蛋白呈递刺突糖蛋白的RBM作为SARS-CoV-2的潜在疫苗姚德辉1、老方1、刘岩1、欧阳方星1、J
。CC-BY-NC-ND 4.0 国际许可,根据 提供(未经同行评审认证)是作者/资助者,他已授予 bioRxiv 永久展示预印本的许可。它是此预印本的版权持有者,此版本于 2020 年 5 月 26 日发布。;https://doi.org/10.1101/2020.05.25.115618 doi:bioRxiv 预印本

以人工智能为核心技术的沿海船舶操舵辅助系统开发
日本正在进入超老龄化社会,近期国内海员存在短缺的风险。此外,由于许多海上事故都是由人为错误引起的,因此强烈需要先进的机械支持来减少船员劳动并提高安全性。在此背景下,船舶操作自动化的趋势正在加速。但要实现这一目标,防止与其他船舶碰撞或搁浅的避让操作必须实现自动化,而这需要超越风险评估,采取规避行动。

横田机场召开飞行安全促进会议(1)
11月2日,第374空运联队安全部邀请约80名当地航空专业人士参加每两年在横田空军基地举办一次的“飞机空中相撞预防措施会议”。该会议于 2010 年首次举办,旨在促进航空安全并加强与当地的双边关系。 (照片 1)11 月 2 日,第 374 维修中队瞬态警报技术员、高级飞行员 Zachary Page 正在指导 MACA 会议参与者驾驶的民用飞机。当地航空官员乘坐商用飞机前往参加 MACA 会议。

通用 DNA 适体可有效抑制 Spike 蛋白/hACE2 相互作用
严重急性呼吸综合征冠状病毒 2 (SARS-CoV-2) 使用其刺突蛋白通过人血管紧张素转换酶 2 (hACE2) 附着到宿主细胞上。可以通过设计一种可以阻断刺突蛋白和 hACE2 之间相互作用的抑制剂来阻止病毒感染。如图 1 所示,一个刺突蛋白三聚体包含三个刺突蛋白,每个刺突蛋白由亚基 S1 和 S2 组成。S1 由 S1A 和 S1B 组成(图 1A),其中 S1B 也称为受体结合域 (RBD),与 hACE2 建立直接相互作用。1 此外,S2 亚基在介导病毒膜与宿主细胞融合方面发挥作用。因此,病毒进入是通过一系列事件完成的,即 S1 与 hACE2 结合,然后触发 S2 将其构象改变为更稳定的融合后状态并允许病毒进入宿主细胞。 2–4 由于 S1 直接与 hACE2 相互作用,许多研究小组一直在积极致力于发现各种生物分子,如抗体 5–10 或适体 11–16,以有效阻断 S1 和 hACE2 之间的相互作用。
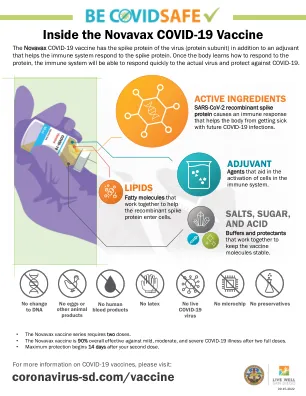
Novavax COVID-19 疫苗内部
Novavax COVID-19 疫苗含有病毒的刺突蛋白(蛋白质亚基),以及帮助免疫系统对刺突蛋白作出反应的佐剂。一旦人体学会如何对蛋白质作出反应,免疫系统将能够快速对实际病毒作出反应并预防 COVID-19。

Notch信号在调节神经元中起双重作用
神经祖细胞会产生兴奋性神经元,其次是少突胶质细胞(OLS)和垂体细胞。然而,调节该神经元时间 - 胶质开关的特定机制尚未完全了解。在这项研究中,我们表明,在胚胎发育的后期阶段,需要在背前祖细胞中Notch信号的适当平衡才能产生少突胶质细胞。在两性的小鼠胚胎中使用离体和子宫方法中,我们发现Notch抑制减少了背胸膜中少突胶质细胞的数量。然而,缺口过度活化也阻止了寡构成并保持祖细胞状态。这些结果表明,在促进和抑制寡头生成中,Notch信号传导的双重作用,必须对其进行微调才能在正确的时间和正确的数字中生成少突胶质细胞谱系细胞。在此过程中,我们进一步将其下游的典型档位hes1和hes5确定为负调节剂。crispr(群集定期间隔短的短质体重复)/cas9介导的hes1和hes5的敲低敲低导致促寡胶质细胞因子ASCL1的表达增加,并导致早产性寡构成。相反,将缺口与ASCL1过表达结合起来,可稳健地促进寡头生成,表明与ASCL1合成的Notch机制单独的机制,以指定少突胶质细胞的命运。我们提出了一个模型,其中Notch信号与ASCL1一起工作以指定祖细胞朝向少突胶质细胞谱系,但也通过hES依赖ASCL1的抑制来维持祖细胞状态,从而使少突胶质细胞不太早,从而导致神经元的精确时间促成神经元 - Glia Switch。

在流行病期间测量疫苗的有效性
COVID-19 疫情带来的紧迫性促使人们实施了适应特殊情况的临床试验 [ 1 ],并建立了史无前例的公私合作伙伴关系 [ 2 ]。特别是,我们观察到,不同疫苗报告的有效性存在很大差异 [ 3 ],从辉瑞和 Moderna(基于 mRNA)的 * 95% 到阿斯利康的 * 70% 或强生的 * 66%(基于病毒载体)。虽然基于 mRNA 的疫苗(如辉瑞-BioNTech 和 Moderna 疫苗)直接指示我们的细胞产生刺突蛋白,但阿斯利康和强生等病毒载体疫苗采用无害的病毒载体将产生刺突蛋白的遗传指令传递给我们的细胞。一旦产生刺突蛋白,我们的免疫系统就会将它们识别为外来物并产生反应。这包括产生可以结合并中和刺突蛋白的抗体,以及激活 T 细胞。这些免疫反应提供了对病毒的免疫力。鉴于所有这些疫苗都是通过刺激刺突蛋白的产生来诱导免疫反应,值得考虑的是,结果的差异是否可能受到实验条件差异的影响,例如感染者的比例和变异的存在。在本文中,我们展示了一些研究结果,这些研究结果表明,在流行病发展的不同阶段测量疫苗效力可能导致对效力的严重低估。

covid-19 - 疫苗成分
也称为表达 SARS-CoV-2 刺突蛋白的 ad26 载体。这是疫苗中唯一的活性成分。这种无害的病毒含有指导人体如何制造可触发免疫反应的刺突蛋白的指令。免疫反应会促使我们产生抗体,如果我们接触到 COVID-19,这些抗体将保护我们。

C 到 U RNA 脱氨是加速 SARS-CoV-2 进化的驱动力
了解SARS-CoV-2猖獗突变的分子机制将有助于我们控制COVID-19大流行。APOBEC介导的C-to-U脱氨是SARS-CoV-2基因组中的主要突变类型。然而,尚不清楚C-to-U的新型突变率u是否高于其他突变类型,以及详细的驱动力是什么。通过分析SARS-CoV-2全球人口数据的时间过程,我们发现C-to-U在所有突变类型中具有最高的新型突变率u,并且该u仍在随时间增加(du / dt> 0)。与其他突变类型相比,新型C-to-U事件对特定的基因组区域具有偏好。局部性较差的RNA结构与较高的新型C-to-U突变率相关。级联模型很好地解释了C-to-U脱氨的du / dt> 0。在SARS-CoV-2中,RNA结构是C到U脱氨速率极高且持续加速的分子基础。该机制是SARS-CoV-2突变、适应和进化的驱动力。我们的发现有助于我们理解病毒突变率的动态演变。