XiaoMi-AI文件搜索系统
World File Search System胞质

使用多能干细胞(例如IPS细胞)进行研究的法规...
建议2.2.1:“鉴于人类胚胎文化的进步以及此类研究的潜力提供有益的发现以促进人类的健康和福祉,ISSCR呼吁国家学院,学术社会,研究授予机构和监管机构与社会有关的社会和社会挑战,并在社会中领导社会挑战,并允许在社会中进行社会的挑战,并允许在社会中进行社会挑战。专业的科学和道德监视过程可以检查14天以上的文化是否是必要的,并且在这种情况下,必须保证用于实现研究目标的胚胎的数量

分子生物学系
1。中村。您的宪法在三年内发生变化。 Shueisha Shinsho,2023年。(第205页)2。中村。环境和表观基因组 - 身体会根据环境而变化吗? - 。 Maruzen Publishing,2018年。(第192)3。中村。表观遗传学,标准分子细胞生物学(印刷),Igakushoin,2024。4。Hino Shinjiro。黄素依赖性组蛋白脱甲基酶的脂肪细胞调节,棕色脂肪组织,CMC Publishing,117-122,2024。5。Hino Shinjiro。通过乳酸代谢,肝胆道胰腺癌重新编程胆管癌(特殊特征:从微环境中解释的胆道胰腺癌),88(5):613-617,2024。6。eto kan,中田Mitsuyoshi。 RNASEQCHEF:自动分析基因表达波动的Web工具,实验医学,41:2307-2313,2023。7。中村。通过代谢和表观基因组控制细胞衰老的机制,生物科学(增强新陈代谢的特殊特征),74:480-481,2023。8。Hino Yuko,Hino Shinjiro,Nakao Mitsuyoshi。通过从线粒体到细胞核的逆行信号的增强剂重塑,医学进度,286:171-172,2023。9。中村。与生活方式有关的疾病:脂肪组织和骨骼肌中的两个代谢表观基因组。途径,饮食和医学,24:21-29,2023。10。Hino Shinjiro。核黄素和黄素蛋白的细胞调节,实验医学补充剂(营养和代谢物信号和食物功能),40(7):1161-1167,2022。11。KOGA TOMOSHO,Nakao Mitsuyoshi。转录组和表观基因组的综合分析,遗传分析新技术及其应用,Wako Pure Chemical Times,89:10-11,2021。 12。 Hino Shinjiro,Araki Yuki,Nakao Mitsuyoshi。肥胖的环境反应敏感的表观基因组形成和个体差异,实验医学特别版(肥胖研究以了解个体差异),5:139-144,2021。 13。 Hino Shinjiro。营养环境适应中的表观遗传学控制机制,基本老化研究,45(3):19-24,2021。 14。 Araki Yuki,Hino Shinjiro,Nakao Mitsuyoshi。表观基因组介导的营养感应和维护和代谢稳态,糖尿病和内分泌代谢部,51:315-322,2020。 15。 Anan Kotaro,Nakao Mitsuyoshi。小儿遗传疾病和表观遗传学,遗传医学穆克独立体积(最新的遗传医学研究和遗传咨询),医学DO,48-53,2019。 16。 中村。健康与疾病(DOHAD)和表观遗传学的发展起源,早产儿,如何成长和发育低流血儿童 - 从出生到Aya一代 - 东京Igakusha,198-208,2019。 17。 Anan Kotaro,Hino Shinjiro,Nakao Mitsuyoshi。组蛋白脱甲基LSD1对骨骼肌细胞的代谢重编程,生物化学,91:31-37,2019。 18。 中村。你和我为什么与众不同?物种与遗传科学,日本临床营养协会杂志,34:19-23,2018。KOGA TOMOSHO,Nakao Mitsuyoshi。转录组和表观基因组的综合分析,遗传分析新技术及其应用,Wako Pure Chemical Times,89:10-11,2021。12。Hino Shinjiro,Araki Yuki,Nakao Mitsuyoshi。肥胖的环境反应敏感的表观基因组形成和个体差异,实验医学特别版(肥胖研究以了解个体差异),5:139-144,2021。13。Hino Shinjiro。营养环境适应中的表观遗传学控制机制,基本老化研究,45(3):19-24,2021。14。Araki Yuki,Hino Shinjiro,Nakao Mitsuyoshi。表观基因组介导的营养感应和维护和代谢稳态,糖尿病和内分泌代谢部,51:315-322,2020。15。Anan Kotaro,Nakao Mitsuyoshi。小儿遗传疾病和表观遗传学,遗传医学穆克独立体积(最新的遗传医学研究和遗传咨询),医学DO,48-53,2019。 16。 中村。健康与疾病(DOHAD)和表观遗传学的发展起源,早产儿,如何成长和发育低流血儿童 - 从出生到Aya一代 - 东京Igakusha,198-208,2019。 17。 Anan Kotaro,Hino Shinjiro,Nakao Mitsuyoshi。组蛋白脱甲基LSD1对骨骼肌细胞的代谢重编程,生物化学,91:31-37,2019。 18。 中村。你和我为什么与众不同?物种与遗传科学,日本临床营养协会杂志,34:19-23,2018。Anan Kotaro,Nakao Mitsuyoshi。小儿遗传疾病和表观遗传学,遗传医学穆克独立体积(最新的遗传医学研究和遗传咨询),医学DO,48-53,2019。16。中村。健康与疾病(DOHAD)和表观遗传学的发展起源,早产儿,如何成长和发育低流血儿童 - 从出生到Aya一代 - 东京Igakusha,198-208,2019。17。Anan Kotaro,Hino Shinjiro,Nakao Mitsuyoshi。组蛋白脱甲基LSD1对骨骼肌细胞的代谢重编程,生物化学,91:31-37,2019。 18。 中村。你和我为什么与众不同?物种与遗传科学,日本临床营养协会杂志,34:19-23,2018。Anan Kotaro,Hino Shinjiro,Nakao Mitsuyoshi。组蛋白脱甲基LSD1对骨骼肌细胞的代谢重编程,生物化学,91:31-37,2019。18。中村。你和我为什么与众不同?物种与遗传科学,日本临床营养协会杂志,34:19-23,2018。
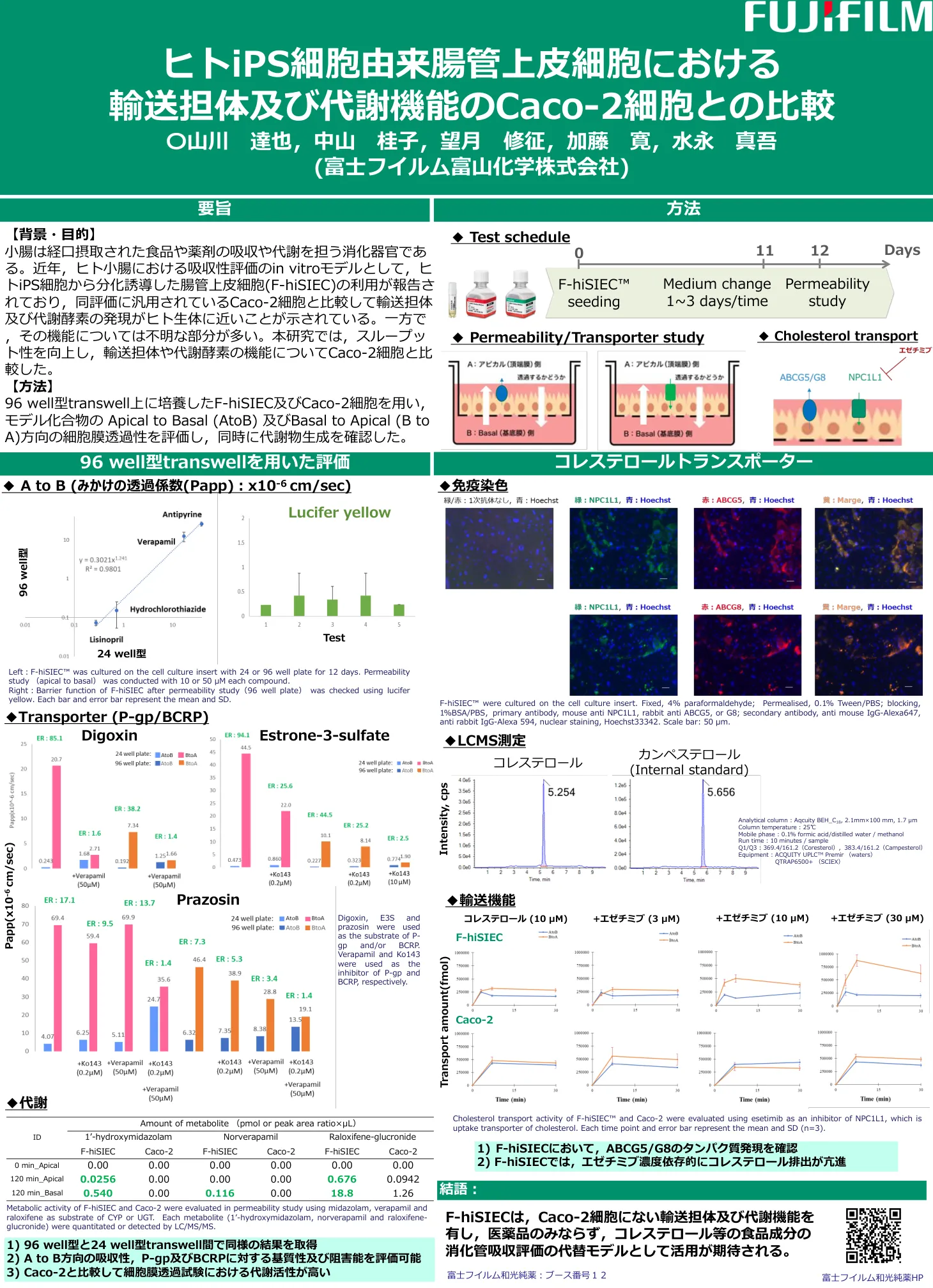
人类 iPS 细胞衍生的肠上皮细胞中的转运蛋白和代谢...
[背景与目的] 小肠是负责口服食物和药物的吸收和代谢的消化器官。近年来,有报道称利用由人类iPS细胞分化而来的肠上皮细胞(F-hiSIEC)作为评价人体小肠吸收情况的体外模型,结果显示其转运载体和代谢酶的表达比通常用于该评价的Caco-2细胞更接近人体。然而,其功能的许多方面仍然未知。本研究提高了通量,并将运输载体和代谢酶的功能与Caco-2细胞进行了比较。 [方法] 利用在96孔Transwell中培养的F-hiSIEC和Caco-2细胞,评估了模型化合物从顶端到基底(A到B)和从基底到顶端(B到A)方向的细胞膜通透性,并同时确认了代谢物的产生。

胞质DNA积累通过独立于STING的途径促进乳腺癌免疫原性
抽象背景免疫检查点阻滞(ICB)已彻底改变了癌症治疗。但是,仅ICB仅在一小部分乳腺癌患者中显示出受益。最近的研究表明,靶向DNA损伤反应的药物可以提高ICB的功效并促进胞质DNA积累。然而,最近的临床试验表明,这些药物与血液学毒性有关。迫切需要更有效的治疗策略。使用多重免疫组织化学染色,将原发性三重阴性乳腺癌肿瘤染色用于胞质单链DNA(ssDNA)。为了增加胞质ssDNA,我们遗传沉默的Trex1。使用鼠类乳腺癌模型评估了肿瘤胞质囊在促进肿瘤免疫原性和抗肿瘤免疫反应中的作用。结果,我们发现肿瘤性胞质ssDNA与乳腺癌三重阴性乳腺癌患者的肿瘤淋巴细胞相关。TREX1缺乏通过DDX3X触发了与STING无关的先天免疫反应。由于Trex1缺失引起的肿瘤中的胞质ssDNA积累足以大大提高ICB的疗效。我们进一步确定了胞质ssDNA诱导剂CEP-701,该溶剂将乳腺肿瘤敏感到ICB,而没有与抑制DNA损伤反应有关的毒性。结论这项工作表明,胞质ssDNA积累促进了乳腺癌的免疫原性,并且可能是一种新型的治疗策略,可以提高ICB毒性的疗效。

使用人类干细胞产生的生殖细胞的人类胚胎...
Murakami K,Hamazaki N,Hamada N,Nagamatsu G,Okamoto I,Ohta H,Nosaka Y,Semba Y,Hayashi K.在体外雄性小鼠的功能性卵母细胞的产生。 div>自然。 div>2023年3月; 615(7954):900-906。 doi:10.1038 / s41586-023-05834-x。 div>


来自各种哺乳动物的多能干细胞的生殖细胞...
1,Hyashi K,Ohta H,Kurimoto K,Saitou M(2011111111 11通过多能干细胞在培养中对小鼠生殖细胞指定途径的重构。单元格,146,519 -2, Gafni O, Weinberger L, Mansour AA, Manor YS, Chom- sky E, Ben-Yosef D, Kalma Y, Viukov S, Maza I, Zviran A, Rais Y, Shipony Z, Mukamel Z, Krupalnik V, Zerbib M, Geula S, Caspi I, Schneir D, Shwartz T, Gilad S, Amann-Zalcenstein D,Benjamin S,Amit I,Tanay A,More-S-RWA R,Novershtern N,Hanna JH(2013年,新型人类基态幼稚的多能干细胞的脱颖而出。 自然,504,282 - 3,Irie N,Weinberger L,Tang WWC,Kobayashi T,Viukov S,Manor YS,Dietmann S,Hanna JH,Surani MA(2015 17是人类原始LOM LOM细胞脂肪的关键特征。 单元格,160,253 - 4, Sasaki k, yokayashi s, namurara t, okamoto i, yabot y, kurimoto k, ohta H, moritoki y, iwatani C, tsuciya h, nakura s, sekiguchi k, sakuma t, yamomomomomoto t, t, yamamoto s, yamamoto m, yamamoto m。 M((2015年)在体外耐心诱导人类生殖细胞脂肪中的人类生殖细胞脂肪。 细胞干细胞,17,178 - 5,Kobayashi T,Zhang H,Tang WWC,Irie N,Withey S,Klipsch D,Syrirna,Dietmann S,Contreras,Webb R,Erlelegio R,Ellelegio R,Soup MA (2 自然,546,416 - 6,Tang WWC,Castillo-Venzor A,Gruhn WH,Kobayashi T,Penfold CA,Morgan MD,Sun D,Irie N,Surani MA (20222222222222222,Sequeential Enlancer State reamoulines Remoulines remoulines hu-man enferine hu-man enterline能力和指定。 nat Cell Biol,24,448 -2, Gafni O, Weinberger L, Mansour AA, Manor YS, Chom- sky E, Ben-Yosef D, Kalma Y, Viukov S, Maza I, Zviran A, Rais Y, Shipony Z, Mukamel Z, Krupalnik V, Zerbib M, Geula S, Caspi I, Schneir D, Shwartz T, Gilad S, Amann-Zalcenstein D,Benjamin S,Amit I,Tanay A,More-S-RWA R,Novershtern N,Hanna JH(2013年,新型人类基态幼稚的多能干细胞的脱颖而出。自然,504,282 -3,Irie N,Weinberger L,Tang WWC,Kobayashi T,Viukov S,Manor YS,Dietmann S,Hanna JH,Surani MA(2015 17是人类原始LOM LOM细胞脂肪的关键特征。 单元格,160,253 - 4, Sasaki k, yokayashi s, namurara t, okamoto i, yabot y, kurimoto k, ohta H, moritoki y, iwatani C, tsuciya h, nakura s, sekiguchi k, sakuma t, yamomomomomoto t, t, yamamoto s, yamamoto m, yamamoto m。 M((2015年)在体外耐心诱导人类生殖细胞脂肪中的人类生殖细胞脂肪。 细胞干细胞,17,178 - 5,Kobayashi T,Zhang H,Tang WWC,Irie N,Withey S,Klipsch D,Syrirna,Dietmann S,Contreras,Webb R,Erlelegio R,Ellelegio R,Soup MA (2 自然,546,416 - 6,Tang WWC,Castillo-Venzor A,Gruhn WH,Kobayashi T,Penfold CA,Morgan MD,Sun D,Irie N,Surani MA (20222222222222222,Sequeential Enlancer State reamoulines Remoulines remoulines hu-man enferine hu-man enterline能力和指定。 nat Cell Biol,24,448 -3,Irie N,Weinberger L,Tang WWC,Kobayashi T,Viukov S,Manor YS,Dietmann S,Hanna JH,Surani MA(2015 17是人类原始LOM LOM细胞脂肪的关键特征。单元格,160,253 -4, Sasaki k, yokayashi s, namurara t, okamoto i, yabot y, kurimoto k, ohta H, moritoki y, iwatani C, tsuciya h, nakura s, sekiguchi k, sakuma t, yamomomomomoto t, t, yamamoto s, yamamoto m, yamamoto m。 M((2015年)在体外耐心诱导人类生殖细胞脂肪中的人类生殖细胞脂肪。 细胞干细胞,17,178 - 5,Kobayashi T,Zhang H,Tang WWC,Irie N,Withey S,Klipsch D,Syrirna,Dietmann S,Contreras,Webb R,Erlelegio R,Ellelegio R,Soup MA (2 自然,546,416 - 6,Tang WWC,Castillo-Venzor A,Gruhn WH,Kobayashi T,Penfold CA,Morgan MD,Sun D,Irie N,Surani MA (20222222222222222,Sequeential Enlancer State reamoulines Remoulines remoulines hu-man enferine hu-man enterline能力和指定。 nat Cell Biol,24,448 -4, Sasaki k, yokayashi s, namurara t, okamoto i, yabot y, kurimoto k, ohta H, moritoki y, iwatani C, tsuciya h, nakura s, sekiguchi k, sakuma t, yamomomomomoto t, t, yamamoto s, yamamoto m, yamamoto m。 M((2015年)在体外耐心诱导人类生殖细胞脂肪中的人类生殖细胞脂肪。细胞干细胞,17,178 -5,Kobayashi T,Zhang H,Tang WWC,Irie N,Withey S,Klipsch D,Syrirna,Dietmann S,Contreras,Webb R,Erlelegio R,Ellelegio R,Soup MA (2自然,546,416 -6,Tang WWC,Castillo-Venzor A,Gruhn WH,Kobayashi T,Penfold CA,Morgan MD,Sun D,Irie N,Surani MA (20222222222222222,Sequeential Enlancer State reamoulines Remoulines remoulines hu-man enferine hu-man enterline能力和指定。nat Cell Biol,24,448 -7,Yamashiro C,Sasaki K,Yabuta Y,Kojima Y,成熟T,Okamoto I,Yokayashi S,Murase Y,Shirara Y,Shirane K,Sasaki K,Sasaki H,Sasaki H,Yamamoto T,Yamamoto T,Saitou M( 201818年)Pluripot pluripot pluripot pluripot celped pluripot pluripot cel celed pluripot pluripot celed pluripot celed细胞的pluripot卷成pluripot。科学,362,356 -8,Hwang YS,Suzuki S,Seita Y,ITTO J,Sa Sato Y,Dog Y,Sato K,Sato K,Hermann BP,Sasaki K (2020020020重建了繁荣症状的spefiification in Verrom,该spefiification in Verrom a Verrom受到了诱发的PACECACE PAMAPOPOTENT SPOS SPOS细胞。nat commun,11,Kobayashi T,Kobayashi H,Goto T,Takashima T,Oakawa M,Ikeda H,Terada R,Yoshida F,Sanbo M,Ukida H,Kurrimoto K,Hirabayashi M (2020 U 2020 U型生殖器开发Kobayashi T,Kobayashi H,Goto T,Takashima T,Oakawa M,Ikeda H,Terada R,Yoshida F,Sanbo M,Ukida H,Kurrimoto K,Hirabayashi M (2020 U 2020 U型生殖器开发

开发脊椎病变性治疗更新*
1。Grupp SA,Kalos M,Barrett D等。嵌合抗原回收的T细胞,用于急性淋巴白血病。n Engl J Med。2013; 368:1509 - 1518。2。Maude SL,Frey N,Shaw PA等。嵌合抗原受体T细胞,可持续减轻白血病。n Engl J Med。2014; 371:1507 - 1517。3。Wherry EJ。 t细胞耗尽。 nat免疫。 2011; 12:492 - 499。 4。 Wherry EJ,Ha SJ,Kaech SM等。 在慢性病毒感染期间CD8 +T细胞耗尽的分子特征。 侵害。 2007; 27:670 - 684。 5。 Zajac AJ,Blattman JN,Murali-Krishna K等。 由于活化的T细胞的持续存在而没有效应子功能,病毒免疫逃避。 J Exp Med。 1998; 188:2205 - 2213。 6。 Martinez GJ,Pereira RM,Aijo T等。 转录因子NFAT促进了激活的CD8 (+)T细胞的耗尽。 免疫。 2015; 42:265 - 278。 7。 Blackburn SD,Shin H,Freeman GJ,Wherry EJ。 通过alphapd-l1阻断对耗尽的CD8 T细胞子集选择性扩展。 Proc Natl Acad Sci U S A. 2008; 105:15016 - 15021。 8。 Pauken KE,Sammons MA,Odorizzi PM等。 表观遗传sta-Wherry EJ。t细胞耗尽。nat免疫。2011; 12:492 - 499。4。Wherry EJ,Ha SJ,Kaech SM等。在慢性病毒感染期间CD8 +T细胞耗尽的分子特征。侵害。2007; 27:670 - 684。 5。 Zajac AJ,Blattman JN,Murali-Krishna K等。 由于活化的T细胞的持续存在而没有效应子功能,病毒免疫逃避。 J Exp Med。 1998; 188:2205 - 2213。 6。 Martinez GJ,Pereira RM,Aijo T等。 转录因子NFAT促进了激活的CD8 (+)T细胞的耗尽。 免疫。 2015; 42:265 - 278。 7。 Blackburn SD,Shin H,Freeman GJ,Wherry EJ。 通过alphapd-l1阻断对耗尽的CD8 T细胞子集选择性扩展。 Proc Natl Acad Sci U S A. 2008; 105:15016 - 15021。 8。 Pauken KE,Sammons MA,Odorizzi PM等。 表观遗传sta-2007; 27:670 - 684。5。Zajac AJ,Blattman JN,Murali-Krishna K等。由于活化的T细胞的持续存在而没有效应子功能,病毒免疫逃避。 J Exp Med。 1998; 188:2205 - 2213。 6。 Martinez GJ,Pereira RM,Aijo T等。 转录因子NFAT促进了激活的CD8 (+)T细胞的耗尽。 免疫。 2015; 42:265 - 278。 7。 Blackburn SD,Shin H,Freeman GJ,Wherry EJ。 通过alphapd-l1阻断对耗尽的CD8 T细胞子集选择性扩展。 Proc Natl Acad Sci U S A. 2008; 105:15016 - 15021。 8。 Pauken KE,Sammons MA,Odorizzi PM等。 表观遗传sta-病毒免疫逃避。J Exp Med。1998; 188:2205 - 2213。 6。 Martinez GJ,Pereira RM,Aijo T等。 转录因子NFAT促进了激活的CD8 (+)T细胞的耗尽。 免疫。 2015; 42:265 - 278。 7。 Blackburn SD,Shin H,Freeman GJ,Wherry EJ。 通过alphapd-l1阻断对耗尽的CD8 T细胞子集选择性扩展。 Proc Natl Acad Sci U S A. 2008; 105:15016 - 15021。 8。 Pauken KE,Sammons MA,Odorizzi PM等。 表观遗传sta-1998; 188:2205 - 2213。6。Martinez GJ,Pereira RM,Aijo T等。转录因子NFAT促进了激活的CD8 (+)T细胞的耗尽。免疫。2015; 42:265 - 278。7。Blackburn SD,Shin H,Freeman GJ,Wherry EJ。 通过alphapd-l1阻断对耗尽的CD8 T细胞子集选择性扩展。 Proc Natl Acad Sci U S A. 2008; 105:15016 - 15021。 8。 Pauken KE,Sammons MA,Odorizzi PM等。 表观遗传sta-Blackburn SD,Shin H,Freeman GJ,Wherry EJ。通过alphapd-l1阻断对耗尽的CD8 T细胞子集选择性扩展。Proc Natl Acad Sci U S A.2008; 105:15016 - 15021。 8。 Pauken KE,Sammons MA,Odorizzi PM等。 表观遗传sta-2008; 105:15016 - 15021。8。Pauken KE,Sammons MA,Odorizzi PM等。表观遗传sta-

免疫和细胞治疗新进展学术会议
肿瘤衍生的外泌体在将肿瘤分子转移到相邻细胞中起着关键作用,从而导致其表型改变并促进肿瘤生长,转移,耐药性和调节肿瘤微环境。我们对恶性脑肿瘤的研究表明,胶质母细胞瘤干细胞(GSC)通过细胞外囊泡(EVS)将其货物转移到肿瘤非茎细胞或正常细胞中,从而导致具有治疗性耐药性和癌性特性的肿瘤干细胞亚型的发展。tzab-001,一种由GSC衍生的细胞外囊泡产生的单克隆抗体,通过阻断EV的细胞间传播,可以显着降低肿瘤干细胞的治疗性耐药性。TZAB-001在神经胶质瘤中识别的蛋白质比其他肿瘤高了60倍。免疫组织化学染色和蛋白质印迹表明,TZAB-001抗体明确识别人GBM干细胞,肝癌细胞系HEPG2,胰腺癌细胞PANC-1和肺癌细胞系A549,但不是正常脑细胞。结果表明,TZAB-001具有肿瘤诊断,癌症干细胞的CART免疫疗法以及ADC药物开发的应用潜力,以提高其功效。

探讨医师在未来「再生细胞治疗」的角色
近年大家对外泌体(外泌体)治疗疾病的相治疗疾病的相:甚至有些学者把异体的外泌:首先外泌体的萃取非常困难复:首先外泌体的萃取非常困难复,理论上必须要把长满干细胞盒子内的培养液,理论上必须要把长满干细胞盒子内的培养液,放在冷冻超高速离心机,放在冷冻超高速离心机10万转,超过,超过12个小时以上,(50-200nm),所以可以穿过所以可以穿过(血脑屏障,Bbb)已,这应该叫msc的条件培养基。至于,经由身体的需求,让干细胞在身体的微环境内