XiaoMi-AI文件搜索系统
World File Search System超螺旋

真核SMC蛋白在每个DNA处诱导-0.6的扭曲...
真核生物携带三种类型的结构性维持(SMC)蛋白复合物,冷凝蛋白,粘着素和SMC5/6,它们是ATP依赖性运动蛋白,通过DNA环挤出重塑基因组。SMCS调制DNA超螺旋,但仍未完全了解如何实现这一目标。在这里,我们提出了一个单分子磁性镊子测定法,该测定法直接测量每个回路 - 分解步骤中单个SMC诱导的扭曲程度。我们证明,所有三个SMC复合物都将相同的较大的负扭曲(即,链接数变化δk k k k占-0.6在每个回路 - 排除步骤中)中的挤压循环,与步长大小无关。使用ATP-Hydrolsyssys突变体和不可用的ATP类似物,我们发现ATP结合是ATPase循环期间的扭曲诱导事件,它与产生力的环路 - 分解步骤相吻合。所有三种真核SMC蛋白诱导相同数量的扭曲表明这些SMC复合物中常见的DNA环境解开机制这一事实。

人类 RNA 聚合酶 I 结构揭示了 HMG-...
RNA 聚合酶 (Pol) I 对核糖体 RNA 前体的转录是细胞生长的主要决定因素,并且在许多癌症类型中都观察到了失调。在这里,我们展示了从携带最大亚基上的基因组 GFP 融合的细胞中纯化人类 Pol I,从而可以跨物种进行酶的结构和功能分析。与酵母相反,人类 Pol I 带有单亚基柄,体外转录表明校对活性降低。在接近天然状态下确定人类 Pol I 低温电子显微镜重建可合理化疾病相关突变的影响,并揭示内置于 Pol I 亚基 RPA1 序列中的额外结构域。这个“dock II”结构域类似于无法与 DNA 结合的截短的 HMG 盒,可作为后生动物的下游转录因子结合平台。生化分析、原位建模和 ChIP 数据表明,拓扑异构酶 2a 可通过域被募集到 Pol I,并与包含因子 UBF 的 HMG 盒域协同作用。这些后生动物 Pol I 转录系统的适应性可能允许有效释放在转录泡下游积累的正 DNA 超螺旋。

Norman F. Moody FRSC的传记
诺曼·穆迪(Norman Moody)于10月23日在维多利亚去世,享年88岁,是许多才华的人。他对多伦多大学的贡献是他在45年以上的独特而杰出的科学生涯中所做的贡献。是生物医学电子研究所的创始董事,随后是生物材料与生物医学工程研究所,以及电气工程学教授。所有这些都没有获得任何获得学位。诺曼于1915年12月22日在第一次世界大战的早期出生于英国赫恩湾。在1930年代初期经济萧条的情况下,如果没有进行正规教育的机会,但对无线电设计的兴趣浓厚,他的职业生涯始于一家大型伦敦商店的无线电维修区。在第二次世界大战之前的几年中,贝尔德(Baird)展示了电视的实际可行性,而五极管和超螺旋接收器已经成为现实。在1935年左右,诺曼(Norman)加入了Halcyon Radio最初以初级工程师的身份从事无线电接收器设计工作,并升任高级电视设计师的位置。接下来是为期一年的设计工程师,在电视研究中与BurnDept Radio进行了设计工程师。在早期的电视设计是一种艺术,需要特殊的实用技能,创造性的思想和巡回设计的才能。他的一些早期作品与投影电视计划的设计有关,但真正的面包和黄油在商用电视机的设计和销售中。当时的一个好系统以大约44几内亚的价格出售,相当于1930年代后期的200美元。正是在他与Halcyon Radio期间,他
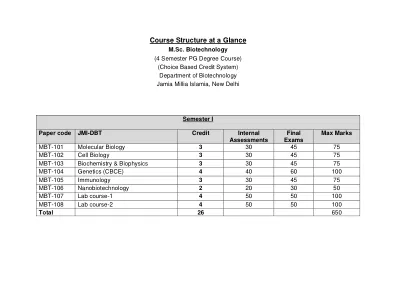
课程结构一览
理学硕士 I 期 MM:75 分子生物学单元 1:基因组的结构和组织(8 小时)染色质组织 - 组蛋白和 DNA 相互作用组、染色质结构、核小体、染色质组织和重塑、染色体、异染色质和真染色质、扭转应力、DNA 拓扑结构 - 链接数、扭曲、扭动、超螺旋、拓扑异构体。第二单元:DNA复制、修复和重组(8 小时)DNA复制模型,Meselson 和 Stahl 实验,DNA聚合酶,病毒、细菌和真核生物中的 DNA 复制,复制叉,复制的校对和保真度,末端复制问题和端粒酶,复制抑制药物,DNA损伤剂,DNA修复机制(核苷酸切除修复、碱基切除修复、错配修复、重组修复、双链断裂修复、转录偶联修复、重组——同源、非同源和位点特异性重组)第三单元:基因表达和调控(8 小时)原核和真核基因的结构、调控区域、转录因子、转录机制、RNA聚合酶、RNA加工结构和不同 RNA 类型的功能、起始复合物的形成、延长、终止;操纵子概念-乳糖操纵子、色氨酸操纵子、arb操纵子、𝜆-阻遏物、lexA阻遏物、噬菌体的溶源性和溶解性循环、核糖开关、转录抑制剂。

分枝杆菌中 CarD 的转录调控受基础启动子动力学指导
细菌病原体,如结核分枝杆菌 ( Mtb ),利用转录因子来使其生理适应宿主内的不同环境。 CarD 是一种保守的细菌转录因子,对 Mtb 的生存至关重要。与通过结合特定 DNA 序列基序来识别启动子的传统转录因子不同, CarD 直接与 RNA 聚合酶结合,以在转录起始期间稳定开放复合中间体 (RP o )。我们之前使用 RNA 测序表明,CarD 能够在体内激活和抑制转录。然而,尽管结合任何 DNA 序列,CarD 如何在 Mtb 中实现启动子特异性调控结果仍不清楚。我们提出了一个模型,其中 CarD 的调控结果取决于启动子的基础 RP o 稳定性,并使用来自具有不同 RP o 稳定性水平的一组启动子的体外转录来测试该模型。我们表明,CarD 直接激活 MTB 核糖体 RNA 启动子 rrnA P3 (AP3) 的全长转录本产生,并且 CarD 的转录激活程度与 RP o 稳定性呈负相关。利用 AP3 的延伸 -10 和鉴别器区域中的靶向突变,我们表明 CarD 直接抑制形成相对稳定 RP o 的启动子的转录。DNA 超螺旋也会影响 RP o 稳定性并影响 CarD 调控的方向,这表明 CarD 活性的结果可受启动子序列以外的因素调控。我们的研究结果为 RNA 聚合酶结合转录因子(如 CarD)如何根据启动子的动力学特性发挥特定的调控结果提供了实验证据。

拓扑异构酶 I 抑制剂在治疗间充质恶性肿瘤中的当前作用及其未来作为肉瘤特异性抗体-药物偶联物的有效载荷的潜在用途
摘要背景:拓扑异构酶 I 是一种酶,它通过松弛超螺旋双链 DNA 在 DNA 复制和转录中起着至关重要的作用。拓扑异构酶 I 抑制剂与拓扑异构酶 I 裂解复合物结合,从而稳定它并防止 DNA 链重新连接,导致 DNA 损伤、细胞周期停滞和细胞凋亡。各种拓扑异构酶 I 抑制剂已在实体瘤中得到评估,伊立替康和拓扑替康已被批准用于治疗上皮恶性肿瘤。这些药物均未获准用于治疗肉瘤,肉瘤是一类多样化的罕见实体瘤,对有效治疗的需求尚未得到满足。摘要:拓扑异构酶 I 抑制剂已在临床前研究中作为单一药物或联合药物在实体瘤中得到评估,其中一些研究包括肉瘤,其中观察到了活性。临床试验评估拓扑异构酶 I 抑制剂治疗肉瘤的效果,结果表明其作为单一疗法疗效有限。与其他细胞毒性药物联合使用时,拓扑异构酶 I 抑制剂已成为特定肉瘤亚型的临床常规治疗手段。伊立替康/长春新碱/替莫唑胺等方案用于治疗复发性横纹肌肉瘤,伊立替康/替莫唑胺和长春新碱/拓扑替康/环磷酰胺通常用于治疗难治性尤文氏肉瘤,拓扑替康/卡铂显示出一定活性

以 DNA 拓扑异构酶 I 为靶点,对中药化合物进行计算筛选和分子对接,设计潜在的抗癌药物
每年,全球有成千上万的人因癌症发病率和死亡率上升而受苦。此外,癌症患者的治疗选择也很昂贵,而且抗癌药物往往疗效较低且副作用较大。DNA拓扑异构酶可以作为已确定的癌症靶点,因为人类拓扑异构酶(Top1)在有丝分裂后阶段调节基因转录,并在复制和修复过程中在DNA超螺旋中起关键作用。因此,在药物治疗过程中,阻断Top1可能对抑制癌细胞增殖至关重要。这里,通过虚拟筛选对中药化合物进行了筛选。中药库的虚拟筛选过程使得能够根据结合能(-7.1至-9.3Kcal/mol)将化合物列表缩小到29种化合物,而在Lipniski过滤之后,使用MM/PB(GB)SA过滤来筛选剩下的22种化合物,并根据结合自由能选出前四种化合物。这里,这四种化合物; CID-65752(T2972:吴茱萸次碱)、CID-5271805(T4S2126:银杏黄素)、CID-9817839(T2S2335:脱氢吴茱萸碱)和CID-51106(T3054:达伍里索林)在分子对接过程中的结合能分别为-8.2、-8.5、-8.3和-8.2,高于其他化合物。在这四个化合物中,ADMET筛选未发现两个筛选化合物CID-5271805和CID-9817839的毒性特征。此外,药物-蛋白质复合物的SASA(溶剂可及表面积)、Rg(回转半径)、RMSD(均方根偏差)和RMSF(均方根波动)轮廓在分子动力学模拟研究中揭示了化合物的稳定性和刚性。然而,这些研究需要通过实验方法进行验证,以开发更有效的抗癌药物。
DinG、RecG 和 RecQ 解旋酶的作用
摘要:富含鸟嘌呤的 DNA 可以折叠成高度稳定的四链 DNA 结构,称为 G-四链体 (G4)。它们最初是在端粒和致癌基因启动子的序列中发现的,可以改变 DNA 代谢。事实上,G4 形成序列代表 DNA 聚合酶的障碍,对细胞生命有重要影响,因为它们可能导致基因组不稳定。为了了解它们在细菌基因组不稳定中的作用,将不同的 G-四链体形成重复序列克隆到大肠杆菌遗传系统中,该系统报告了当 G 道在复制过程中包含前导或滞后模板链时重复序列的移码和完全或部分缺失。这些重复序列在单链 DNA 中形成稳定的 G-四链体,但在天然超螺旋双链 DNA 中不形成。尽管如此,转录促进了 (G 3 T) 4 和 (G 3 T) 8 重复序列在所得 R 环中形成 G-四链体。根据遗传背景和序列结构形成的倾向,突变率相差 5 个数量级。此外,虽然体外方法表明细菌解旋酶可以分解 G4,但目前仍不清楚 G4 解旋在体内是否重要。在这里,我们表明 recG 突变会降低突变率,而结构特异性解旋酶 DinG 和 RecQ 的缺陷会增加突变率。这些结果表明 G-四链体的形成会促进细菌的遗传不稳定性,解旋酶在体内控制这一过程中起着重要作用。

方案1- gacose凝胶电泳
GAROSE是一种线性聚合物,由A-(L-73)和糖苷键连接的交替残基和L-半乳糖组成。L-半乳糖残留物具有三个至六个位置之间的避别桥(请参见图5-1)。琼脂糖的链形成螺旋纤维,将半径为20-30 nm的超螺旋结构聚集。琼脂糖的凝胶化会导致三维通道的网格,其直径从50 nm到> 200 nm(Norton等人。1986;有关审查,请参见Kirkpatrick 1990)。 商业制备的琼脂糖聚合物被认为每个链中包含半乳糖残基。 但是,琼脂糖不是均匀的:多糖链的平均长度因批量而异,从制造商到制造商。 此外,琼脂糖的较低等级可能会被其他多糖以及盐和蛋白质污染。 这种变异能力可以影响琼脂糖溶液的胶凝温度,DNA的筛分以及从凝胶中回收的DNA的能力,可作为酶促反应中的底物。 可以使用特殊的琼脂糖等级来最大程度地减少这些潜在的问题,这些琼脂糖被筛选为抑制剂和核酸酶的存在以及用溴化乙锭染色后的最小背景荧光。1986;有关审查,请参见Kirkpatrick 1990)。商业制备的琼脂糖聚合物被认为每个链中包含半乳糖残基。但是,琼脂糖不是均匀的:多糖链的平均长度因批量而异,从制造商到制造商。此外,琼脂糖的较低等级可能会被其他多糖以及盐和蛋白质污染。这种变异能力可以影响琼脂糖溶液的胶凝温度,DNA的筛分以及从凝胶中回收的DNA的能力,可作为酶促反应中的底物。可以使用特殊的琼脂糖等级来最大程度地减少这些潜在的问题,这些琼脂糖被筛选为抑制剂和核酸酶的存在以及用溴化乙锭染色后的最小背景荧光。

教学大纲
ns cc11-(th)-p01:生物分子,酶学和仪器生物分子:生命的化学基础 - 化学键合,涉及生物分子的力和构建块 - 大分子;信息大分子。蛋白质作为信息大分子;氨基酸的化学;多肽的一级,二级和三级结构;肽;肽亚基和第四纪结构, -helix,-薄片和胶原蛋白结构,蛋白质和氨基酸的代谢。碳水化合物的化学 - 单,二糖和多糖。DNA的分子结构,替代DNA结构,圆形和超螺旋DNA,DNA的变性和恢复,DNA的物理和化学稳定性。酶和反应动力学:酶的定义;活性位点,底物,辅酶,辅因子和不同种类的酶抑制剂;酶动力学,两种底物动力学,三种底物动力学,偏离线性动力学;配体结合研究;快速动力学;关联和解离常数;在酶动力学机理分析中使用同位素; pH,温度和同位素标记的底物对酶活性的影响;酶调节的变构模型;底物诱导酶的构象变化。电子显微镜:磁性和静电镜的理论及其焦距;电子显微镜的构造;限制分辨率和有用的放大倍数;对比形成;阴影和染色技术;扫描电子显微镜;标本准备技术;电子显微镜在细胞和分子生物学中的应用;嵌入和切割。仪器:生物系统光谱后的原理和应用:吸收光谱(UV-可见),荧光和磷光,圆形二色性(CD),红外光谱学(IR),共振拉曼光谱;电子旋转共振(ESR),液体闪烁计数器; pH计;超速离心,光学显微镜,光学显微镜;阶段,紫外线和干扰显微镜 - 其基本原理;光学系统和射线图 - 它们在细胞生物学中的应用;荧光显微镜;细胞和组织的微光照射法,荧光活化的细胞分辨率(FACS)。