XiaoMi-AI文件搜索系统
World File Search System酵母

酵母 PHO5 启动子处核小体结构的有效动力学
摘要染色质动力学由重塑酶介导,在基因调控中起着至关重要的作用,正如在典型模型酿酒酵母 PHO5 启动子中建立的那样。然而,有效的核小体动力学,即启动子核小体配置的轨迹,仍然难以捉摸。在这里,我们通过整合已发表的单分子数据推断出这种动力学,这些数据捕获了从受抑制到完全活跃的 PHO5 启动子状态的多核小体配置,以及其他现有的组蛋白周转和新的染色质可及性数据。我们设计并系统地研究了一类新的“受调节的开关滑动”模型,模拟全局和局部核小体(解)组装和滑动。68,145 个模型中只有 7 个与所有数据吻合良好。所有七个模型都涉及滑动和 N-2 核小体的已知核心作用,但通过调节一个组装而不是解体过程来调节启动子状态转换。这与 PHO5 启动子先前观察结果的常见解释一致,但提出了挑战,并表明染色质通过结合竞争而开放。

致病酵母近平滑念珠菌中多态性着丝粒的位置
着丝粒提出了一个进化悖论:功能高度保守,但序列和结构却迅速变化。然而,在没有损伤的情况下,着丝粒的位置通常在一个物种内是保守的。我们在此报告,致病酵母菌种近平滑假丝酵母的分离株在其八条染色体中的两条染色体上表现出着丝粒位置的种内多态性。它的旧着丝粒具有反向重复 (IR) 结构,而其新着丝粒没有明显的结构特征,但位于旧位置的 30 kb 以内。因此,着丝粒可以自然地从一个染色体位置移动到另一个染色体位置,似乎是自发的,并且在 DNA 序列没有任何显著变化的情况下。我们的观察结果与所有着丝粒都是由基因决定的模型相一致,例如由短或长 IR 的存在或形成十字形的能力决定。我们还发现着丝粒已成为 C. parapsilosis 进化枝中基因组重排的热点。

最终合成酵母染色体完成,为生物技术进步铺平道路
该项目的共同负责人、澳大利亚研究理事会合成生物学卓越中心主任、杰出教授伊恩·保尔森 (Ian Paulsen) 表示:“通过成功构建和调试最终的合成染色体,我们帮助完成了一个强大的工程生物学平台,这可能会彻底改变我们生产药品、可持续材料和其他重要资源的方式。”

CRISPR/CAS9用于开发酿酒酵母细胞工厂的系统
合成酵母细胞工厂为一系列产品的可持续供应提供了一个显着的解决方案,从大型工业化学品到高价值药品化合物。合成生物学是一个领域,在其中对代谢途径进行了深入研究和设计。群集,定期间隔,短,全文重复相关(CRISPR)/CRISPR相关蛋白9(CAS9)技术已成为合成生物学的最新基因编辑技术。最近,使用不同的CRISPR/CAS9系统的使用已扩展到酵母工程领域,用于单核苷酸分辨率编辑,多基因编辑,转录调控和基因组规模的修改。这种进步系统导致了涉及减少劳动和时间的加速微生物工程,并增强了对细胞遗传学和生理学的理解。本评论简要概述了最新的研究进度以及CRISPR/CAS9系统在遗传操作中的使用,重点是酿酒酵母酿酒酵母细胞工厂工程的应用。

酵母提取物agar磷酸盐缓冲盐水,pH 7.2,10x
用户必须在使用前确保产品在其应用中的适用性。产品仅符合该和其他相关HIMEDIA™出版物中包含的信息。本出版物中包含的信息基于我们的研发工作,据我们所知,真实而准确。Himedia™实验室Pvt Ltd保留随时更改规格和信息的权利。产品不是用于人类或动物诊断或治疗用途的产品,而是用于实验室,研究或进一步制造的使用,除非另有说明。本文包含的陈述不应被视为任何形式的保证,明示或暗示,也不应对侵犯任何专利的责任承担任何责任。

营养
酵母菌补液(例如SuperStart®)由酵母衍生的生长和生存因子(麦角固醇,长链脂肪酸)配制,其目的是增强酵母膜的元素。superStart®被吸收到酵母细胞膜中,以帮助膜的流动性,酒精和温度耐受性,并具有较高的糖和氮转运蛋白效率。酵母补液产品的好处延伸到最后一代酵母。酵母重新填充产物不含在补液阶段对酵母有毒的DAP。酵母营养素含有生长因子,生存因子和促进酵母菌繁殖。复杂的酵母营养素是用酵母自动材料,灭活酵母,DAP和硫胺素配制的。酵母营养素,以促进健康的酵母生物量,并在必须缺乏氮的情况下支持酵母。使用SuperStart®(酵母再补水产品)和Nutristart®(发酵营养素)将确保健康的发酵,增强酵母衍生的香气产生,并减少VA和H 2 S生产。
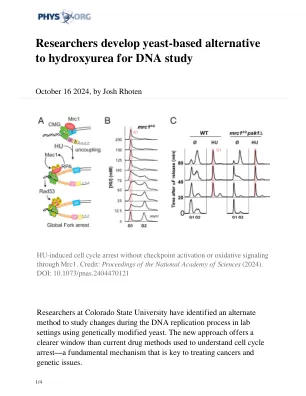
研究人员开发出基于酵母的羟基脲替代品用于 DNA 研究
该研究成果发表在《美国国家科学院院刊》上,由科罗拉多州立大学生物化学和分子生物学系助理教授 Grant Schauer 领导。该研究重点研究羟基脲,这是一种用于临床治疗癌症的化疗药物,也经常用于研究以阻止细胞发育周期。这样做可以让研究人员更好地探索基因组 DNA 在细胞分裂前准确复制的复杂过程。

适应性实验室进化对提升非常规酵母菌种生物技术潜力的贡献
摘要:通过选择压力和全基因组重测序控制实验室中的短期进化过程,可以诱导几代生物特性的变化,这有助于确定微生物适应性实验室进化 (ALE) 的遗传基础。由于该技术的多功能性以及对石油基策略替代品的迫切需求,ALE 已在多种酵母中积极开展,主要使用常规酿酒酵母,但也使用非常规酵母。由于转基因生物是一个有争议的话题,而且全球尚未就其使用达成共识,因此 ALE 成为了当前的热门话题,出现了大量采用 ALE 方法的新研究,并在此背景下开发了许多不同的应用。在本综述中,我们首次收集了相关研究,这些研究展示了非传统酵母物种的 ALE 对其生物技术改进的作用,并根据研究目的对其进行了分类,并根据所用的物种、实验结果和所采用的方法对其进行了比较。本综述阐明了 ALE 作为一种强大工具的适用性,可以增强物种特征并提高其在生物技术中的性能,重点是非传统酵母物种,作为替代方法或与基因组编辑方法相结合。

人类疾病酵母模型中磷光酶缺乏症的进化营救
摘要糖基化(CDG)的人类先天性疾病的最常见原因是磷光合酶基因PMM2中的突变,它影响蛋白质N-连接的糖基化。酵母基因SEC53编码人类PMM2的同源物。我们进化了384个酵母,载有两个与人疾病相关的等位基因之一,SEC53-V238M和SEC53 -F126L或野生型SEC53。我们发现,1000代后,大多数种群弥补了与Sec53人疾病相关等位基因相关的慢增长表型。通过全基因组测序,我们确定了补偿性突变,包括已知的SEC53遗传相互作用。我们观察到其他基因的补偿性突变富集,其人类同源物与1型CDG相关,包括PGM1,该基因编码酵母中磷酸葡萄糖核酶的少量同工型。通过遗传重建,我们表明进化的PGM1突变是主要的,并且是特异性的遗传相互作用者,可恢复具有Sec53 -V238m等位基因的蛋白质糖基化和酵母的生长。最后,我们表征了纯化的PGM1突变蛋白的酶活性。我们发现,PGM1活性的减少(而不是消除)最好地补偿了与Sec53 -V238M等位基因相关的有害表型。广义,我们的结果证明了实验进化的力量,作为识别补偿与人类疾病相关等位基因的基因和途径的工具。

基于 CRISPR 的葡萄酒酵母技术概述,旨在改善葡萄酒的风味和安全性
摘要:现代工业酿酒以使用特定的葡萄酒菌株发酵剂为基础。商业葡萄酒菌株比天然分离物具有多种优势,它们的使用保证了工业酿酒技术的稳定性和可重复性。对于竞争激烈的葡萄酒市场以及对提高葡萄酒质量和葡萄酒安全性的新需求,开发新的酵母菌株变得越来越重要。在过去的几十年里,在实验室中创造升级的葡萄酒酵母的新可能性出现了,从而开发出具有更好发酵能力的菌株,能够改善葡萄酒的感官品质并生产针对特定消费者的葡萄酒,考虑到他们的健康和营养需求。然而,只有两种转基因 (GM) 葡萄酒酵母菌株正式注册并获准用于商业用途。与传统的基因工程方法相比,CRISPR/Cas9 被描述为高效、多功能、廉价、易于使用,并且能够靶向多个位点。该基因工程技术自 2013 年以来已应用于酿酒酵母。在这篇评论中,我们旨在概述 CRISPR/Cas9 编辑技术在葡萄酒酵母中的应用,以结合开发能够增加葡萄酒中风味化合物而不会产生异味的表型,并有助于创造“更安全的葡萄酒”。