XiaoMi-AI文件搜索系统
World File Search System附着的

沿海沉积物的间质微生物群落由纳米章节主导
地下沿海沉积物中的微生物群落高度多样,并且在营养循环中起着重要作用。,虽然沙质沉积物中的微生物的主要部分呈足为Epipsammon(附着在沙粒上),但只有一小部分在间质毛孔中繁殖。到目前为止,对这些自由生活微生物群落的组成知之甚少。在这项研究中进行了研究,在沙滩的地下中,我们比较了沉积物中的古细菌和细菌群落结构,以及应用16S rRNA基因测序的相应毛孔水。我们发现,根据孔隙空间的不同,自由生活原核生物的比例仅为0.2-2.3%。间质微生物群落显示出一个小的重叠,附着的分数为4-7%,并且包含在孔道中仅发现的75-81%ASV的独特组成。它们比各自的沉积物级分更多样化,并且显示出更高的古细菌比。古细菌主要隶属于Dpann Superphylum的纳米章,相对丰富的间隙群落相对丰富。细菌分数包括与候选门辐射(CPR)有关的几种物种。已知两种原核生物谱系都有小细胞尺寸,包括尚未尚未识别的代谢功能的尚未培养的物种。我们的发现得到了对相邻潮汐平坦的调查,显示出类似的趋势。因此,我们的结果表明在沿海沉积物的地下存在不同的间质微生物群落。这种尚未培养的纳米章的自然富集和心肺复苏群的成员为靶向元基因组分析甚至隔离这些群体成员提供了进一步代谢表征的机会。
![arxiv:2209.00266v2 [cond-mat.soft] 2023年5月11日](/simg/9\9f9f0214d9e4581bffe3991608c5eb8d129e69fe.webp)
arxiv:2209.00266v2 [cond-mat.soft] 2023年5月11日
运动蛋白(MP)是真核细胞中cy骨骼的组成部分[1-3]。它们参与了亚细胞过程中的广泛功能,例如货物的细胞内转运,细胞骨架动力学,应力产生和细胞运动。他们水解ATP以经过附着的结局,并沿着附着状态的共轭纤维进行分解运动[4-8]。例如,MPS的动力蛋白和动力蛋白沿微管移动,而MPS的肌球蛋白家族可以沿纤维肌动蛋白移动。他们的运动取决于载荷[9,10],并且他们可以达到的最大ve-受到可用的ATP浓度[11]。ATP水解对化学物质的局部耗散驱动MPS脱离平衡。他们的运动方向取决于可以行走的局部前后不对称性。在最小的尺度上生成非平衡驱动,MP构成了一类活动物质[12-14],其中时间反转对称性和平衡闪烁 - 耗散关系被损坏。在活细胞中,MP共同运输包括细胞器在内的各种货物[15-19]。从几个到数百个国会议员可以参与这种运输[20-25]。多个MP驱动的货物动力学的理论研究使用相等的负载共享近似值或有限数量的MPS的详细数值模拟[26-33]。弹性耦合MPS显示应变诱导的解开和停滞[37 - 39]。除了进行细胞内反式 -MPS之间的耦合可能是由直接的机械连接产生的,如肌球蛋白纤维[34],分子拥挤效应[35,36]或与货物的结合,尚未完全了解其可能的影响。用于弱构层,有效的解开速率和平均货物载体恢复到单运动行为的非相互作用限制。
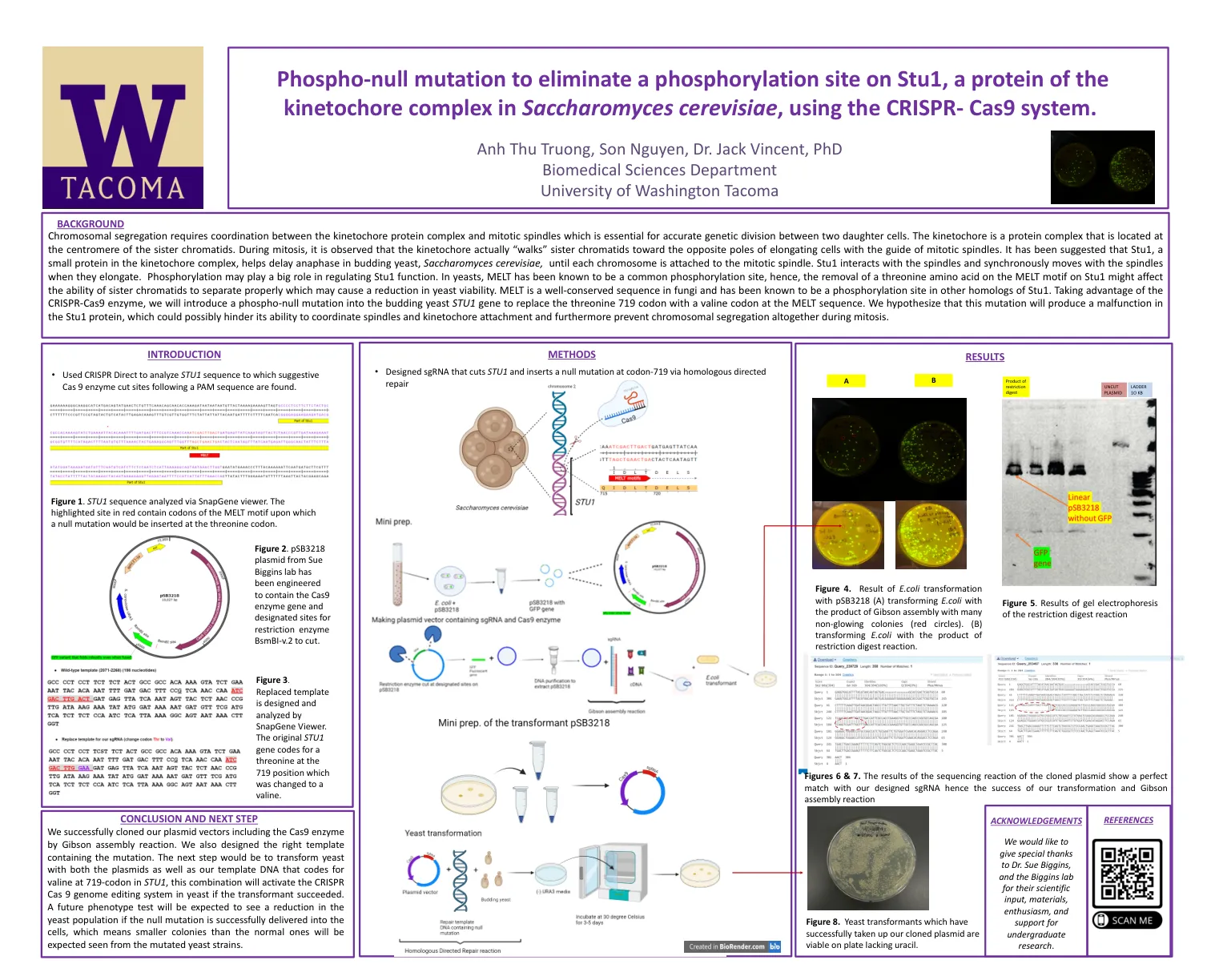
利用 CRISPR 技术进行磷酸化无效突变,消除酿酒酵母动粒复合体蛋白 Stu1 上的磷酸化位点
染色体分离需要动粒蛋白复合物和有丝分裂纺锤体的协调,这对于两个子细胞之间的准确遗传分裂至关重要。动粒是一种位于姊妹染色单体着丝粒的蛋白复合物。在有丝分裂过程中,可以观察到动粒实际上是在有丝分裂纺锤体的引导下将姊妹染色单体“引导”到伸长细胞的相反极点。有人提出,动粒复合物中的小蛋白 Stu1 有助于延迟芽殖酵母酿酒酵母的后期,直到每条染色体都附着在有丝分裂纺锤体上。Stu1 与纺锤体相互作用,并在纺锤体伸长时与其同步移动。磷酸化可能在调节 Stu1 功能方面发挥重要作用。在酵母中,MELT 是一种常见的磷酸化位点,因此,去除 Stu1 上 MELT 基序上的苏氨酸氨基酸可能会影响姐妹染色单体正确分离的能力,从而导致酵母活力下降。MELT 是真菌中保存良好的序列,并且已知是 Stu1 其他同源物中的磷酸化位点。利用 CRISPR-Cas9 酶,我们将在芽殖酵母 STU1 基因中引入磷酸化无效突变,以将 MELT 序列中的苏氨酸 719 密码子替换为缬氨酸密码子。我们假设这种突变会导致 Stu1 蛋白发生故障,这可能会阻碍其协调纺锤体和着丝粒附着的能力,并进一步阻止有丝分裂期间染色体分离。

建模由地中海中生物型微塑料介导的碳导出
海洋微塑料可以通过生物污染的微生物生物定植,从而导致微塑料的浮力降低。因此,生物质塑料的下沉可以代表海洋碳循环中新型的碳出口途径。在这里,我们建模了微塑料如何通过杜型生物融合,由于浮力变化而导致的垂直运动以及水柱中粒子附着的硅藻和碳池之间的相互作用。我们使用来自Nemo-Medusa-2.0的生物地球化学数据初始化了Lagrangian框架,并估算出以100 mM微塑料的不同表面浓度从1 mm微塑料的不同表面浓度开始的有机碳的量。我们专注于以世界上一些最高的微塑料浓度为特征的Medi-TerraneA海,并且是由大气中二氧化碳水平上升引起的生物地球化学变化的热点。我们的结果表明,下沉的生物融合微塑料引起的碳输出与海面层中的微塑料浓度成正比,至少在建模浓度下。我们估计,尽管当前的微塑料浓度可以使自然生物碳的导出<1%,但未来在业务上的污染场景下预测的未来浓度可能会导致碳出口量超过基线(1998 - 2012年),到2050年。以高主要生产力为特征的区域,即西地中海和中部,是微塑料介导的碳出口结果最高的地区。虽然强调了这种现象在地中海中的潜力和数量有限的发生,但我们的结果呼吁进一步研究全球海洋中与微塑料相关的碳出口途径。

在L模式高通力运动期间,西部主要等离子体面向组件的钨大侵蚀调查
在2023年,在西部进行了最初的高通力运动,该活动是由新安装的主动冷却的钨分流,由Iter级单块组成。该活动包括在附着的转移条件下重复60秒钟的长氘L模式脉冲,累积了超过10000秒的血浆暴露。在外部罢工点区域达到了大约5举10 26 m -2的最大氘静脉,代表了一些高性能iTer脉冲。从可见光谱中推断出的总钨侵蚀表明,最受侵蚀的等离子体面向成分是内部分流目标,其速率比外移分离目标大十倍。位于离等离子体数厘米的外部平面钨保险杠,显示出侵蚀率的侵蚀速率是外移分流的两倍。我们得出的结论是,外部平面保险杠对远程钨的迁移和沉积到下层的延长具有可忽略的贡献。内部分流器上的累积总侵蚀率以约20μm的有效总侵蚀厚度转换,而外分离器的侵蚀速率约为20μm。引人注目的是,这些订单与分流物上本地的沉积物厚度一致:高场侧单块的裸露表面覆盖着几个μm的钨沉积物,而在下部侧面,很少有μm薄钨沉积物仅在磁性阴影部分上发现单块的磁性阴影部分。尤其是研究行动的开始,应考虑分离侵蚀预算的定义,以预测有害存款的形成。这些沉积物对西部运行的强烈影响,即表面温度测量与红外热摄影的扰动以及片的发射导致受限血浆的辐射扰动,要求预测ITER中的类似问题。

ran2a低MOA提出了Fonsi ...
拟议中未根据《国家环境政策法》(NEPA)的规定(42美国法典(U.S.C.)§§4321-4347);环境质量委员会(CEQ)NEPA法规(40条联邦法规[CFR]第1500-1508部分);空军(DAF)环境影响分析过程(EIAP)(32 CFR第989部分)和联邦航空管理局(FAA)命令(FAA)命令1050.1F,环境影响:政策和程序,DAF准备了附加的环境评估(EA),以评估拟议中的潜在环境后果,从而从拟议中的行动中进行了较低的行动,以建立一个新的Persorent Senonement Saniore san New-Al-al-al-al-al-al-al-alt al-al-al-al-al-al-al-al-al-al-alt al-alt al-al-al-al-al-al-al-al-al-alt al-al-al-al-al-altertimile(Moiore)(Moiore)(Moiore)。拟议的行动将支持在圣安东尼奥 - 兰多夫空军基地联合基地(JBSA-Randolph),德克萨斯州以及其他暂时性DAF用户的第12飞行训练部(12 ftw)的未来未来试点培训要求。根据国防部与FAA之间的理解备忘录,FAA正在作为合作机构作为合作机构,以根据FAA订单JO 7400.2p的特殊用途空域(SUA)诉讼的环境审查,处理空间问题。作为此过程的一部分,FAA将与航空用户公开通行拟议的领空,以征求信息,以帮助确定它对可通道的空域产生什么影响。除了与此EA相关的公众参与外,还将发生循环化。在FAA循环过程中收到的评论将在最终的EA中考虑,如适用。DAF将要求FAA发行自己的fonsi,并在DAF发出fonsi后尽快将RAN2A低MOA领空图表。附着的EA是通过参考将其纳入的,该发现没有显着影响(FONSI)。

su(3)扭曲的梯度流强耦合无...
摘要:由于人为活性,海洋的汞含量(HG)含量增加了两倍,尽管黑海洋(> 200 m)已成为重要的HG储层,但有毒和生物蓄积的甲基汞(MEHG)的浓度很低,因此很难测量。因此,当前对深海中HG周期的理解受到严格的数据限制,控制MEHG的因素及其转换率仍然很大程度上未知。通过分析52个全球分布的巴基拉质深元素宏基因组和26个来自Malaspina Expedition的新元转录组,我们的研究揭示了在全球浴类海洋中(〜4000 m深度)中细菌编码基因Mera和Merb的广泛分布和表达。这些基因与Hg II还原和MEHG脱甲基化相关的基因在粒子附着的分数中尤为普遍。此外,我们的结果表明,水质量年龄和有机物组成塑造了拥有Mera和Merb基因的结构,这些群落和Merb基因生活在不同的粒径分数,其丰度及其表达水平。命令的成员Corynebacteriales,Rhodobacterales,Alteromonadales,Oceanospirillales,Moraxelleles和Flavobacteriales是深海中包含Mera和Merb基因的主要分类参与者。这些发现,加上我们先前具有具有代谢能力降解MEHG的深层层流海洋的纯培养物分离株的结果,表明甲基汞脱甲基化和HG II还原可能发生在全球黑暗海洋中,这是生物圈中最大的生物组。关键字:汞,甲基汞,浴样,细菌脱甲基化,宏基因组,metatranscriptomes,mer基因■简介

社论:益生菌研究中的OMICS技术和生物信息学工具
益生菌被定义为活的微生物,可以促进肠道和肠外健康的好处,当时有足够的数量消耗(Hill等,2014)。由于其安全性和促进健康的特性,几种双杆菌,乳酸杆菌和肠球菌已被分类为益生菌。这些微生物通常在各种栖息地中发现,例如乳制品和非乳制发酵产物,哺乳动物胃肠道菌群和环境。为了将新的菌株分类为益生菌,应满足许多标准:抗胃肠道转移的抵抗力,缺乏毒力和可传播的抗生素耐药性基因以及促进健康的活性(例如抗菌,免疫抑制性和抗毒剂和抗毒剂)。监管机构已经建立了常规的微生物学测定,以评估这些表型(FAO/WHO,2001)。此外,现在正在使用高通量多词方法来补充现有方法,并将更深层次的分子和细胞见解与益生菌 - 宿主相互作用(Kiousi等,2021)。在(元)基因组学时代,益生菌菌株的整个基因组序列(WGS)的可用性呈指数增长。基因组元素在益生菌研究中的整合支持了新菌株的安全性和功能性的预测。此外,由于其较高的歧视能力,WGS是将新分离株分类为物种分类分类的“黄金标准”。的确,WGS的可用性增加促进了多样化的乳杆菌属的重新分类。基于共同的生态和代谢特性,分为25属(Zheng等,2020)。目前,EFSA需要在食物链中使用微生物WG,以监测关注的基因(例如,毒力因子,抗生素耐药性基因)(EFSA,2024年)。在这种情况下,Wei等人进行了补充了体外测定的基因组分析。评估limosilactobacillus reuteri A51的安全性和功能性状,这是先前从Yak酸奶中分离出来的菌株。菌株被发现编码与胃肠道应力反应,生存和附着的基因以及用于抗菌化合物和外多糖的生物合成簇。该菌株还表现出对模拟胃肠道条件以及抗氧化剂和

使用CRISPR
染色体隔离需要在动型蛋白复合物和有丝分裂纺锤体之间进行协调,这对于两个子细胞之间的遗传分裂至关重要。动力学是一种蛋白质复合物,位于姐妹染色单体的丝粒上。在有丝分裂过程中,观察到的动物学实际上将姐妹染色质朝着用有丝分裂纺锤体的指南伸向细胞的相反两极。有人提出,stu1是一种小动物络合物中的小蛋白,有助于延迟酿酒酵母的萌芽酵母中的后期,直到每个染色体都附着在有丝分裂的纺锤体上。也有人建议Stu1与纺锤体相互作用,并在拉长时同步移动。已经提出,磷酸化可以调节Stu1的功能,并且熔体是其他动力学蛋白中已知的磷酸化位点,因此,在称为sTu1上的称为熔融基序的磷酸化位点上除去苏氨酸氨基酸在Stu1上的磷酸化位点可能会影响姐妹染色体的能力,这可能会导致姐姐的正确性,这可能会使YEAST YEAST降低。熔体是真菌中保存良好的序列,是其他动力学蛋白中的已知磷酸化位点,是STU1的同源物。利用CRISPR-CAS9酶,我们将在发芽的酵母菌Stu1基因中引入磷酸无效突变,以用熔体序列替代苏氨酸719密码子。到目前为止,我们已经成功克隆了含有引导RNA和Cas9酶基因的质粒。我们假设该突变将在Stu1中产生故障,这可能会阻碍其协调纺锤体和动孔附着的能力,并在有丝分裂过程中完全防止染色体分离。下一步将是用质粒和我们的模板DNA转化酵母,该模板DNA代码在Stu1中的719密码子上编码Valine,这种组合将完全激活酵母中的CRISPR CAS CAS 9基因组编辑系统。

自然风格的仿生表面,用于控制细菌附着和生物膜发育
估计,由于抗生素耐药细菌引起的感染每年享有70万寿命。如果耐药细菌继续以相同的速度进化,则预计到2050年,该数字将增加到1000万。[5]同样,生物和非生物表面上生物膜的形成对人类健康也面临着另一个重大挑战。生物膜形成。[2,3,6]在生物膜形成期间,微生物经历了几种生物学变化,导致细胞外聚合物物质(EPS)产生。EPS矩阵可防止并从攻击生物膜的化学物质和其他毒素中产生微生物。这使得很难使用传统的抗生素治疗生物膜,并使微生物获得抗菌抗性。[7]解决此问题的一种方法是使用抗菌材料和表面,以抑制抑制性细胞的附着和生物膜的形成。[4,8–13]这种抗菌材料作为工程材料的作用至关重要,因为这些材料可能有助于我们减少对抗生素和消毒剂的依赖。已经采用了各种技术来制造抗臭材料,包括用杀菌层涂上材料表面。[11,14–18]常见方法是基于从材料表面(例如金属衍生物和抗生素)释放杀菌剂的。[24]因此,当地形特征的尺寸在亚微米和纳米长度尺度中时,表面会抑制细菌的附着。[18]一种替代方法涉及将微/纳米摄影应用于消除细菌细胞的应用,并且在过去十年中,这种方法引起了很大的兴趣。[19-21]具有独特的微/纳米尺度表面纹理和特征的材料已被证明可以有效禁止细菌附着并防止生物膜形成。[22,23]提出,当地形特征的尺寸小于细菌细胞的尺寸时,降低了附着的细胞的可用接触区域。由于其表面地形特征,许多天然材料已经获得了防染色和反抗性行为。[11,25,26]这激发了科学家开发类似的材料和结构,这些材料和结构限制了生物膜形成并积极消除与表面接触的细菌。这样的