XiaoMi-AI文件搜索系统
World File Search System鞭毛藻

S12915_020_00927_9.pdf
背景:鞭毛藻是水生生物的人,在全球海洋中特别广泛。有些人负责有毒的花朵,而另一些人生活在共生关系中,既可以作为珊瑚中的共生式共生体,要么是感染其他生物和动物的寄生虫。鞭毛菌具有非典型的大基因组(〜3至250 GB),其基因组织和基因表达模式与密切相关的Apicomplexan寄生虫截然不同。在这里,我们测序并分析了两种早期差异和同时发生的寄生虫鞭毛蛋白变形虫菌株的基因组,以阐明这种非典型基因组特征,鞭毛藻酸酯的进化和宿主专业化的出现。结果:我们使用Illumina配对的短读和牛津纳米孔技术(ONT)的长读测序方法的组合,对两种变形虫菌株(A25和A120)进行了测序,组装和注释的高质量基因组(A25和A120)。我们发现了少数可转座元素,以及短的内含子和基因间区域以及有限的基因家族,共同促进了大变形虫基因组的紧凑性,这一特征可能与寄生虫有关。大多数变形虫蛋白(A25的63.7%和A120的59.3%)没有功能分配,但我们发现许多与Dinophyceae共享的直系同源物。我们的分析表明,尽管种间蛋白质序列相似性低,但两种基因组之间的单向簇编码和高水平的同步保护的基因趋势很强。

diatom nitzschia captiva的遗传转化方法
内共生症在海洋环境中是一种广泛且在生态上具有重要意义的现象。这些内共生伙伴如何发展为具有新细胞器的有机体,仍然是未知的,需要调查现代共生体。dinotoms,具有进化中间硅质质体的dinoflagellates,被认为是研究细胞器发生的出色模型,因为它们在连续三个但不同的阶段保持了。努力了解宿主的鞭毛藻 - 结合硅藻的关系受到了共生的任何一个成员的遗传转化方法的限制。为了解决这种缺陷,我们修改了现有的硅藻生物和共轭转化方法和硅藻nitzschia captiva的冷冻保存方案,这是kleptoplastic dinotom dinotom dinotom dinotom dinotom duinotom durinskia capensis的重要猎物。Through the use of Phaeo dactylum tricornutum, Cylindrotheca fusiformis, and native Nitzschia captiva diatom designed plasmids, we suc cessfully express and target EGFP to the cytosol, mitochondria, and plastids of N. captiva, and visualize these organelles inside D. capensis in vivo, allowing specific labeling and tracking of摄入后的细胞器和蛋白质。此外,我们尝试利用CRISPR/CAS9来瞄准引入的EGFP基因,但找不到成功基因编辑的证据。

水生环境中的微生物多样性
在水生生态系统的水下是一个充满生命的微观宇宙,在维持这些环境的微妙平衡中起着至关重要的作用。水生微生物学探讨了各种水体中微生物的多样性和功能,从广阔的海洋到最小的淡水池塘。在水生环境中,最丰富,最多样化的微生物群是营养循环的关键参与者。例如,硝基瘤和硝化细菌参与硝化过程,将氨转化为氮气中的硝酸盐。一些细菌也有助于有机物的降解,在营养回收中起重要作用。从微观浮游植物到较大的宏观形式,藻类是带有光合作用的阳光的主要生产者。硅藻,鞭毛藻和绿藻是水生食物网的重要贡献者,通过生产有机化合物为各种生物提供了能量。这些单细胞真核生物是水生生态系统中重要的消费者。鞭毛,纤毛和变形虫在调节细菌种群,回收养分以及作为较高营养水平的食物方面起着作用。病毒虽然不是严格归类为生物体,但在水生环境中很丰富,并影响微生物种群。噬菌体,感染细菌的病毒可以调节细菌群落,影响养分循环和微生物多样性。水生微生物对于包括碳,氮和磷循环在内的营养循环过程至关重要。细菌和藻类有助于释放有机物的细分,从而释放出其他生物可以利用的营养。藻类和蓝细菌进行光合作用,将阳光转化为化学能。这个过程不仅支持这些微生物的生长,而且还为其他水生的能源提供了主要的能量
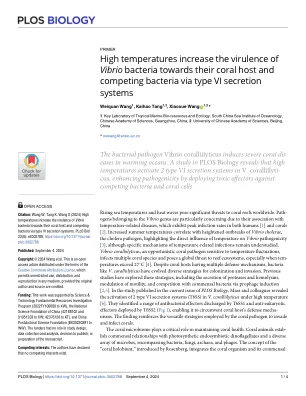
高温增加了通过VI型分泌系统
海上温度和热浪的上升对全世界的珊瑚礁构成了重大威胁。属于弧菌属的途径尤其是由于它们与温度相关疾病的关联,后者在人类和珊瑚[1]和珊瑚[2]中均表现出峰值感染率。夏季温度的升高与霍乱病原体的弧菌病原体的爆发爆发相关,突出了温度对弧菌致病性的直接影响[3],尽管与温度相关感染的特定机制仍然被忽略了。弧菌Coralliilyticus是对温度波动敏感的机会性珊瑚病原体,感染多种珊瑚种类,并对礁生态系统构成全球威胁,尤其是当温度超过27°C时[4]。尽管珊瑚宿主具有多种防御机制,但细菌如V。Coralliilyticus发展了殖民和入侵的多种策略。先前的研究已经探索了这些策略,包括蛋白酶和血素蛋白的分泌,运动能力的调节以及通过预言诱导与共生细菌的竞争[2,5]。在发表在《 PLOS生物学》中发表的研究中,Mass及其同事揭示了V中2型VI型分泌系统(T6SS)的激活。在高温下[6]。他们确定了由T6SS1和T6SS2部署的抗核效应器排放的一系列抗菌效应器(图1),使其能够绕开珊瑚宿主的防御机制。这一发现加强了珊瑚病原体侵入和感染珊瑚的多功能策略。珊瑚微生物组在维持珊瑚健康中起着至关重要的作用。珊瑚动物与光合性内共生鞭毛藻和各种微生物,包括细菌,真菌,古细菌和噬菌体的多种微生物。罗森伯格(Rosenberg

卵菌的转化系统、基因沉默和基因编辑技术
卵菌是一类多样化的丝状产孢生物,由数百种臭名昭著的病原体组成。其中一些已被列入全球检疫名单,并受到国家和国际法律的严格管制,以防止其传播(Rossmann 等人,2021 年)。宿主包括主要养殖鱼类和植物物种,以及自然生态系统中的众多动物和植物物种(Cao 等人,2012 年;Fern andez-Ben eitez 等人,2008 年;Kamoun 等人,2015 年;van den Berg 等人,2013 年)。卵菌是一类在分类学上截然不同的真核微生物大类,它与真菌有一些相同的生理和形态特征(例如,都有菌丝和不同的孢子类型),但在系统发育上与异鞭毛藻有亲缘关系(Baldauf 等人,2000 年;Latijnhouwers 等人,2003 年)。卵菌与真真菌可通过一些只有卵菌才具备的生化和细胞学特征来区分:a) 纤维素是菌丝壁的主要微纤维成分;b) 胞质致密体/指纹液泡含有磷酸化的 β-(1,3)-mycolaminarin 葡聚糖;c) 二倍体叶状体,减数分裂先于配子形成;d) 线粒体有管状嵴;最后 e) 利用 a - ε -二氨基庚二酸赖氨酸合成途径 ( Beakes 等人,2012 年)。卵菌生长的环境条件和宿主范围广泛,这反映在其系统发育多样性中 ( Thines,2014 年)。在过去的几十年里,宿主与卵菌相互作用的研究结合基因组学和转录组学,对卵菌如何感染宿主有了相当深入的了解 ( Burra 等人,2017 年)。了解许多相互作用分子的作用对于有针对性地制定管理策略非常重要。已确定卵菌会分泌一系列效应蛋白,这些效应蛋白可以改变宿主的免疫系统以促进感染(Bozkurt 等人,2012 年;de Bruijn 等人,2012 年;Fabro 等人,2011 年)。然而,不同卵菌病原体在感染过程中产生的大量分子尚未得到解释。为了在体内对这些分子进行功能分析,对卵菌进行基因改造的技术至关重要,例如 RNAi(Saraiva 等人,2014 年;Whisson 等人,2005 年)、稳定转化(Judelson 等人,1993 年)或 CRISPR/Cas(Fang 和 Tyler,2016 年)。卵菌的分子技术发展速度比真菌慢,目前仅限于相对较少的物种,与真菌相比效率较低。由于卵菌内部的异质性,转化方案需要针对每个物种进行优化,并且在同一物种内,通常针对每个菌株进行优化。因此

第一部分 - 阿基默
布雷斯特湾(BB)是法国西北地区的浅河口环境。这个半封闭的盆地为180平方米,受到多种流体动力因素的影响,包括海洋电流和河流放电的双重影响(Aulne和Elorn Main Main Rivers),并导致复杂的水力气候和水力沉积过程。这项研究使用pa孢子数据(大陆:花粉颗粒和海洋:鞭毛藻囊肿)进行研究两种不同的材料:(i)在整个BB上收集的现代表面沉积物以及(ii)三个新的BB沉积物核心(来自Aulne River和Cores Palm-ks05和Palm-ks05和Palm-Krare的口中的三个新的BB沉积物核心(来自Aulne River and Palm-kerreart oft the Brest of the Brest of the Brest of Brest of Brest of Brest of Brest of Brest of。While modern data are analysed from a statistical point of view to highlight the influence of hydrodynamic forcing on the modern distribution of palynomorphs, the cores allow for spatial comparisons of palynological data on three windows over the Early (~9.5 and ~8.5 ka BP), Middle (~4.4–4.3 ka BP interval) and Late (~1–0.9 ka BP interval) Holocene.在每个时间间隔中,比较了从西部(更明显的海洋影响)到东部的两个岩心(比较明显的海洋影响)(来自奥尔恩河的更强烈的河流影响),位于极限的两侧,我们称为河流诱导的元水学信号(RIPS)极限。这些不同的比较表明,随着时间的推移,BB花粉记录中的空间均匀性很高,除了裂口以东的环境外,降雨引起的河流排放的影响更大,尤其是考虑到河岸分类群(即alnus)。这旨在提高对不同BB芯场记录的载流信号的理解,这是在建立覆盖BB不同浅层层中几个核心的多个核心的全新世的孢粉堆栈之前至关重要的相关性的第一步(请参阅Valero等人(参见Valero等人),请参阅Valero等人,分组 - 第二部分)。

淡水藻类示例
藻类品种包括海藻,池塘浮渣和海带都来自同一个家庭。这些生物的植物样特征如叶绿体,可以进行光合作用的LIK植物。有些藻类还鞭毛和中心藻,在饲料习惯方面,它们与动物更相似。藻类范围从微小的单细胞生物到大型多细胞类型,它们生活在各种环境中,包括盐水,淡水,湿土或潮湿的岩石。较大的藻类物种通常被称为简单的水生植物。硅藻是盐水环境中最丰富的浮游生物类型,人数超过金棕色藻类。没有细胞壁,硅藻具有称为浮雕的二氧化硅壳,其形状和结构取决于物种。金棕色藻类虽然不太常见,但被称为纳米膨胀,仅由50微米的细胞组成。消防藻类,也称为鞭毛藻,是单细胞的,当它们大量盛开时会引起红潮,在海洋中以红色的色调出现。某些吡咯烷物种是生物发光的,导致水在夜间发光。鞭毛藻是有毒的,会产生可破坏人和其他生物体肌肉功能的神经毒素。与鞭毛藻类似的Cryptomonads也可能会产生有害的藻华,将水变深褐色或红色。netrium desmid是在淡水和盐水环境中发现的单细胞绿藻类的顺序,在具有对称结构的长丝状菌落中生长。绿藻主要居住在淡水中,但也可以在海洋中找到。F.E.它们具有由纤维素制成的细胞壁,并含有叶绿体,使它们可以进行光合作用。多细胞种类的绿藻形成菌落,从四个细胞到几千个细胞。用于繁殖,一些物种与一个鞭毛一起游泳的非运动型植物孢子或Zoospores。绿藻类的类型包括海莴苣,马毛藻和死者的手指。红藻通常在热带海洋位置发现,生长在珊瑚礁等实心表面或附着在其他藻类上。它们的细胞壁由纤维素和各种碳水化合物组成。红藻通过产生由水流携带的单孢子直至发芽的单孢子。他们还经历了有性繁殖和几代人的交替。不同种类的红藻形成不同的海藻类型,例如以其优雅的外观而闻名的plumaria elegans。海带是在水下海带森林中发现的一种棕色藻类。棕色藻类是最大的藻类类型之一,由在海洋环境中发现的各种海藻和海带组成。它们具有分化的组织,包括锚固器官,浮力的空气口袋,茎,光合器官以及产生孢子和配子的生殖组织。棕色藻类的生命周期涉及世代的交替。一些棕色藻类的例子包括萨尔加苏姆杂草,岩藻和巨型海带,它们的长度最高可达100米。黄绿色藻类是藻类的最少种类的类型,只有几百种,它们是单细胞生物,具有由纤维素和二氧化硅制成的细胞壁。藻类是具有类似于植物的特征的生物。它们最常见于水生环境中,藻类有七种主要类型,每个藻类具有不同的特征。绿藻通常生活在淡水中,而红绿色藻类则生活在新鲜和盐水环境中。本文解释了藻类的不同类型,包括它们的独特特征和栖息地。它还讨论了藻类作为包含植物样特征并具有光合作用的生物的重要性。藻类的大小差异很大,范围从单细胞到大型多细胞物种,并且可以在不同的水生环境以及潮湿的表面上找到。与较高的植物不同,它们没有根,茎,叶或花朵,并且缺乏血管组织。藻类作为主要生产者在水生生态系统中起着至关重要的作用,它是盐水虾和磷虾等各种海洋生物的食物来源。他们通过性和无性恋方法繁殖,一些物种经历了世代的交替。繁殖方法通常取决于温度,盐度和营养供应性等环境因素。Fritsch分类藻类基于色素沉着,thallus结构,储备食品,鞭毛和繁殖方式。藻类的两种主要类型是叶绿素(绿藻)和Phaeophyceae(棕色藻类)。叶绿素科包括约7,000种,主要在具有海洋形式的淡水环境中发现。他们通过性,无性和营养方法繁殖。它们表现出各种结构,例如单细胞,殖民地,丝状和管状形式。绿藻由于含有不同颜料的叶绿体而能够进行光合作用。它们的颜色范围从黄绿色到深绿色,它们具有线粒体,带有平坦的Cristae,中央液泡和由纤维素和果胶制成的细胞壁。Phaeophyceae由大约2,000种生活在海洋环境中。它们的特征是由于高水平的岩甘氨酸而引起的棕色着色,这是诸如Chl-A,C,Carotenes和Xanthophylls之类的光合色素的另一种存在。他们的植物体被分为固定的锚固,长期存在的stipe,lamina或frond可能是一年。海带或海藻在这一组中是显着的较大形式,其中一些物种达到了相当大的尺寸,例如大环(30-60m),使其成为最大的海洋植物。这些藻类包含由纤维素和藻类等多糖制成的细胞壁,纤维素和藻类酸是一种复杂的多糖,有助于保护它们免受各种环境因素的侵害。棕色藻类包含锚定器官,茎,光合器官以及发展孢子和配子的生殖组织。,他们以拉米那肽和甘露醇的形式保留食物,如在拉米那尼亚,大环,内囊等物种等物种中所见。红色藻类具有植物蛋白酶和植物素色素,使它们的颜色显得红色,尤其是在更深的水域中。这些生物可以由于这些色素而吸收蓝绿色的光谱,从而使它们在更大的深度繁殖。一个例子是液泡。大多数红藻是光自人营养的,但有一些例外,例如Harveyella,它生活在其他红藻类上。它们的细胞壁由纤维素,果胶和硫酸化植物胶体(如琼脂)组成。红藻中的thallus组织可以从单细胞到类似蕾丝的结构不等。这些生物可以保留食物为佛罗里达淀粉,在Gonyostomum和Chattonella等物种中发现。黄绿色藻类是最少的多产量,只有450-650种。它们主要是单细胞的,具有纤维素 - 硅细胞壁,用于运动的鞭毛以及缺乏某些色素的叶绿体。Xanthophyceae通常形成细胞的小菌落,并具有用于运动的鞭毛。他们将食物保留为脂肪,主要是在具有盐水适应的淡水环境中发现的。他们的性繁殖很少见。菊科是单细胞或殖民地鞭毛物,包括各种类型的球形,衣壳,丝状,丝状,变形虫,质子和实质形式。大约12,000种菊科,主要是居住在淡水环境中,其中一些在盐水栖息地中发现。这些微生物的特征在于诸如叶绿素A,P-胡萝卜素和叶黄素等色素。黄金藻类以脂肪的形式存储能量,很少经历有性繁殖,并产生称为囊肿的专门静息细胞。运动形式具有一两个不同类型的鞭毛:金属丝或鞭打。chrysocapsa,lagynion,ochromonas,chrysamoeba是金藻的例子。例子包括气旋,thalassiosira,Navicula和Nitzschia。接下来,芽孢杆菌科(硅藻)由约12,000至15,000种。这些微生物在显微镜下显示为鼓形细胞,并带有一些形成的链。硅藻以脂肪的形式存储能量,并经历广泛的有性繁殖。它们具有由果胶和二氧化硅组成的硅化细胞壁,存在于淡水,海洋和陆地环境中。隐藻科是单细胞鞭毛形式,约有200种。在光学显微镜下,它们以红色或红色颜色的逗号形细胞出现。Cryptophyceae以淀粉的形式存储能量,具有由纤维素组成的细胞壁,并具有两个不等的鞭毛。罕见的异恋性繁殖发生在这些生物体中,居住在淡水和海洋环境中。例子包括plagioselmis,falcomonas,rhinomonas,teleaulax和chilomonas。Dinophyceae是大约200种的运动单细胞生物。他们的主要色素包括叶绿素a和c,β-胡萝卜素和叶丁香。罕见的异恋性繁殖发生在这些生物中,这些生物主要居住在海洋环境中,但有些存在于淡水中。Dinophyceae以淀粉或脂肪的形式存储能量。例子包括Alexandrium,Dinophysis,Gymnodinium,Peridinium,Polykrikos,Noctiluca,Ceratium和Gonyaulax。叶绿素科是具有鲜绿色色谱和过量叶丁香的单细胞生物。他们以脂肪的形式存储能量,并具有双足动动物形式。这些微生物仅居住在淡水环境中。euglenineae是具有光合色素的运动单细胞或殖民地生物,例如叶绿素a和b,β-胡萝卜素和木蛋黄酱。他们以淀粉或脂肪的形式存储能量,并具有类似于微观动物的裸纤毛生殖器官。有性繁殖尚未得到这些生物的明确证明。尤格伦氨酸中不存在细胞壁,其中一种或多种金属丝类型。一个例子是Euglena。最后,蓝藻科或粘菌科(蓝绿色藻类)由单细胞,殖民地或多细胞体组成,具有原核核和双膜性线粒体和叶绿体。这些微生物居住在各种环境中,并具有多种特征。颜料在蓝藻科的独特蓝色中起着至关重要的作用,植物蛋白蛋白是主要的贡献者。这组藻类缺乏运动阶段,而以氰基雄雄或粘菌糖淀粉的形式存储食物。它们的细胞壁由果胶或纤维素组成。在许多蓝绿色藻类物种中常见的独特特征,例如“假”分支和杂环。在蓝菌科中没有有性繁殖,无处不在,到处都可以找到。这些生物的例子包括Nostoc,振荡器,Anabaena,Lyngbya和Plectonema。藻类是主要生产者,利用叶绿素A和B进行光合作用,并且具有确定其颜色的各种色素。藻类通常被错误地考虑到植物或生物。然而,某些物种可以产生有毒的花朵,例如红潮,蓝绿色藻类和蓝细菌,对人类健康,水生生态系统和经济构成重大威胁。藻类有多种类型的藻类,包括绿藻(绿藻),Phaeophyceae(棕色藻类),rohodophyceae(红藻类),Xanthophyceae(黄绿色藻类)和氰基藻科和粘液菌科或粘粒细菌(蓝绿色藻类)。这些生物可以大致分为三个大藻类:棕色藻类,绿藻和红藻。