XiaoMi-AI文件搜索系统
World File Search SystemABA

2023 年 ABA 国际法分会 (ILS) 年度会议
该专家学者和从业人员小组将分享他们对国际仲裁最新重大发展的看法,包括审查美国最高法院和英国、加拿大等其他司法管辖区的重大判决。讨论的主题包括美国最高法院在 ZF Automotive 和 AlixPartners 案中的裁决,该裁决限制了联邦地区法院在国际仲裁程序中下令取证的权力,以及在 Morgan v. Sundance 案中的裁决,该裁决裁定,联邦航空管理局不授权联邦法院在裁定一方放弃仲裁权之前要求裁定存在偏见。本课程提供专家对国际仲裁最新法律发展的看法,这将引起所有在该领域工作或对该领域感兴趣的从业人员的极大兴趣。

CRISPR/Cas9 介导的脱落酸受体基因多重敲除降低了大豆种子发芽过程中对脱落酸的敏感性
摘要:脱落酸(ABA)是一种重要的植物激素,参与调节植物生长、发育和逆境响应中的多种功能。多种蛋白质参与调控环境胁迫下ABA信号转导机制,其中PYR1/PYL/RCAR家族为ABA受体。本研究利用CRISPR/Cas9基因编辑系统和单个gRNA敲除大豆三个PYL基因:GmPYL17、GmPYL18和GmPYL19。T0代植株基因分型结果显示,gRNA可有效敲除GmPYL17、GmPYL18和GmPYL19基因靶序列,并使其发生不同程度的缺失。一组诱导的等位基因被成功转移到后代。在T2代,我们获得了双重和三重突变的基因型。在种子萌发阶段,CRISPR/Cas9技术制备的GmPYL基因敲除突变体,尤其是gmpyl17/19双突变体对脱落酸的敏感性低于野生型。利用RNA-Seq技术,通过3个生物学重复研究不同处理下萌发幼苗对脱落酸反应相关的差异表达基因。gmpyl17/19-1双突变体种子萌发过程中对脱落酸的敏感性降低,突变株高和分枝数高于野生型。在脱落酸胁迫下,GO富集分析显示一些正向萌发调控因子被激活,降低了脱落酸敏感性,促进了种子萌发。本研究为从分子水平上深入研究脱落酸信号通路及其关键成分的参与提供了理论基础,有助于提高大豆对非生物胁迫的耐受性,同时也有助于育种者调控和提高大豆在不同胁迫条件下的产量和品质。

激光熔覆(定向能量沉积)中的粉末收集效率。对标准激光熔覆和 ABA 熔覆技术的研究
本文研究了吹粉激光熔覆(一种定向能量沉积技术)中的粉末收集效率。对标准的“逐道重叠”熔覆(“ AAA ”熔覆)和“ ABA ”熔覆进行了比较,其中初始一组间距较大的轨道(“ A ”轨道)之间留下的间隙由后续的“ B ”轨道填充。在这两种技术中,熔池表面是熔覆粉末的收集区域,并且该池的形状会受到多种参数的影响,包括熔覆速度、轨道间距和熔覆技术类型。这里给出的结果来自于对加工过程中拍摄的高速视频和所得熔覆轨道的横截面的分析。结果表明,AAA 熔覆中的第一个轨道与后续轨道具有不同的熔池形状,并且后续轨道熔池的不对称导致粉末收集效率降低。与此相反,“B”轨道熔池与其相邻的“A”轨道之间的几何形状可提高粉末收集效率。
![水稻种子休眠4同源物RDO5 1的逆转与bHLH57相互作用控制拟南芥脱落酸的生物合成和种子休眠[打开]](/simg/4\4bdcb63afc05cf222922d655b34e57eda1e5494c.webp)
水稻种子休眠4同源物RDO5 1的逆转与bHLH57相互作用控制拟南芥脱落酸的生物合成和种子休眠[打开]
脱落酸 (ABA) 对种子休眠的控制已得到广泛研究,但其潜在机制尚未完全了解。本文,我们报告了拟南芥 (Arabidopsis thaliana) 中两种与 ABA 相关的种子休眠调节剂的特征:ODR1(用于逆转 rdo5),水稻 (Oryza sativa) 种子休眠 4 (Sdr4) 的直系同源物,以及碱性螺旋-环-螺旋转录因子 bHLH57。ODR1 的转录水平直接受到转录因子 ABA INSENSITIVE3 (ABI3) 的抑制,它通过影响 ABA 生物合成和 ABA 信号传导来负向调节种子休眠。相比之下,bHLH57 通过诱导基因 9-CIS-EPOXYCAROTENOID DIOXYGENASE6 ( NCED6 ) 和 NCED9 的表达来正向调节种子休眠,这两个基因编码 ABA 生物合成酶,从而导致更高的 ABA 水平。ODR1 与 bHLH57 相互作用并抑制 bHLH57 调节的 NCED6 和 NCED9 在细胞核中的表达。bhlh57 功能丧失等位基因可以部分抵消 odr1 突变体中增强的 NCED6 和 NCED9 表达,因此可以挽救它们相关的超休眠表型。因此,我们确定了一个新颖的 ABI3-ODR1-bHLH57-NCED6/9 网络,该网络为了解 ABA 生物合成和信号传导对种子休眠的调节提供了见解。
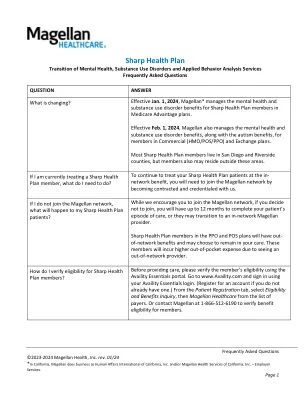
敏锐的健康计划 - 提供商门户 - 麦哲伦医疗保健
所有ABA服务都需要预先授权。要请求初始服务,请填写ABA初始授权表,并通过加密电子邮件将其与会员的诊断报告一起提交给sharphpaba@magellanhealth.com。对于所有随后的请求,请提交您的更新治疗计划或通过加密电子邮件持续护理的Magellan ABA治疗计划/并发审查表格sharphpaba@magellanhealth.com。查找有关自闭症谱系障碍的其他信息,包括索取ABA初始授权和并发审查的表格,请访问www.magellanprovider.com/autism(登录必修)。
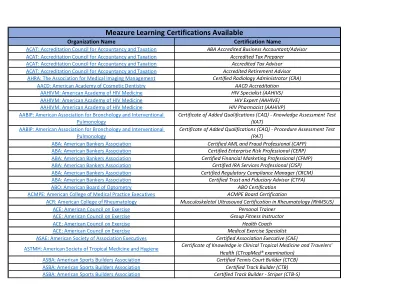
Meazure学习认证
(PAT) ABA: American Bankers Association Certified AML and Fraud Professional (CAFP) ABA: American Bankers Association Certified Enterprise Risk Professional (CERP) ABA: American Bankers Association Certified Financial Marketing Professional (CFMP) ABA: American Bankers Association Certified IRA Services Professional (CISP) ABA: American Bankers Association Certified Regulatory Compliance Manager (CRCM) ABA: American Bankers Association Certified Trust and Fiduciary Advisor (CTFA)ABO:美国验光委员会ABO认证ACMPE:美国医学学院高管ACMPE ACMPE委员会认证ACR ACR ACR:美国风湿病学院骨骼骨骼骨骼骨骼骨骼超声检查(RHMSUS)ACE ACE ACE ACE ACE:ACE)ACE:美国运动委员会运动专家:美国运动委员会:美国运动委员会:美国运动委员会:美国运动委员会:美国运动委员会:美国运动委员会:运动律师运动:运动律师运动:锻炼律师:运动律师:协会主管协会认证协会主管(CAE)

仲裁中的AI和数字化
除了私营部门的创新外,美国律师协会(ABA)等组织还采取了主动性将技术纳入法律实践,包括在替代争议解决方案(ADR)领域。ABA创新中心促进了采用AI和数字工具,以增强司法障碍并提高争议解决过程的效率。ABA还主张在仲裁和调解中使用电子发现工具和虚拟平台,认识到它们降低成本并提高诉讼速度的潜力。此外,ABA正在为制定法律实践中AI使用的道德准则的持续努力帮助解决对AI驱动决策中透明,偏见和问责制的担忧。

自闭症干预的证据、伦理和有效性。
自闭症的主要干预措施之一是应用行为分析 (ABA)。ABA 是一种干预模式,重点是改变自闭症儿童的外部行为,目的是让自闭症儿童看起来和行为起来不像自闭症儿童。自闭症社区内部提出了伦理问题,即 ABA 和类似做法如何通过教孩子隐藏自闭症特征来“正常化”自闭症儿童。隐藏自闭症特征与自闭症成年人的心理健康状况恶化和自杀倾向增加有关。此外,这些做法往往以牺牲促进关键生活技能、自决和自尊为代价来“正常化”自闭症儿童。然而,自闭症社区对自闭症治疗和服务的担忧远远超出了“正常化”问题,甚至超出了 ABA 问题。

应用行为分析亚专业领域
应用行为分析 (ABA) 以其在治疗自闭症谱系障碍 (ASD) 和其他发育障碍(例如唐氏综合症、智力障碍)患者方面的成功而闻名。该领域的治疗对个人的整个生命周期(即童年、青少年、成年)都有效。对于患有自闭症等发育障碍的幼儿,强化、综合干预的目标是提高认知、语言、社交和自助技能。数十年的研究表明,强化 ABA 治疗是治疗自闭症儿童最成功的方法,并得到了美国卫生局局长、美国儿科学会和美国国家心理健康研究所等多个机构的广泛认可。当应用于老年人时,ABA 涉及教授在家庭、学校和社区中有效运作所必需的行为。ABA 还可以减少危害健康和安全的严重问题行为,并限制教育、居住或职业选择。
蜡状芽孢杆菌NJ01通过EDS1-WRKY18模块诱导SA-和ABA介导的对细菌病原体的免疫力
Dacheng Wang, 1 Lirong Wei, 1 Jinbiao Ma, 1 Yingqiao Wan, 1 Keyi Huang, 1 Yiqiong Sun, 1 Huili Wen, 4 Zhipeng Chen, 4 Zijie Li, 1 Dongli Yu, 2 Haitao Cui, 3 Jingni Wu, 1 Yufeng Wu, 4 Sun Tae Kim, 5 Jing Zhao, 1 Jane E. Parker,6 Kenichi Tsuda,7岁, * Chunhao Jiang,1, *和Yiming Wang 1,8, * 1植物病理学系,农作物疾病综合管理和害虫综合管理的主要实验室,Nanjing农业大学教育部Nanjing 210095,NANJING 210095 02115, USA 3 Department of Plant Pathology, College of Plant Protection, Shandong Agricultural University, Tai'an, Shandong 271018, China 4 State Key Laboratory for Crop Genetics and Germplasm Enhancement, Jiangsu Key Laboratory for Information Agriculture, Bioinformatics Center, Academy for Advanced Interdisciplinary Studies, Nanjing Agricultural University, Nanjing, China 5 Department of Plant Bioscience, Life and Industry Convergence Research Institute, Pusan National University, Miryang 50463, Republic of Korea 6 Department of Plant-Microbe Interactions, Max Planck Institute for Plant Breeding Research, 50829 Cologne, Germany 7 State Key Laboratory of Agricultural Microbiology, College of Plant Science and Technology, Huazhong Agricultural University, Wuhan 430070, China 8 Lead contact *Correspondence: tsuda@mail.hzau.edu.cn(K.T.),chjiang@njau.edu.cn(C.J.),ymwang@njau.edu.cn(y.w。)https://doi.org/10.1016/j.celrep.2024.113985