XiaoMi-AI文件搜索系统
World File Search SystemEPAS1

功能性 EPAS1/ HIF2A 错义变异与安第斯高地人的血细胞比容有关
Elijah S. Lawrence 1†,Wanjun Gu 1†,Ryan J. Bohlender 2,Cecilia Anza-Ramirez 3,Amy M. Cole 4,James J. Yu 1,Hao Hu 2,Erica C. Heinrich 1.5,Katie A. O'Brien 1.6,Katie A. O'Brien 1.6,Carlos A. vasquez 7,Quinh kny tickhhy tickhha t. I 1.9,TAO长9.10,James E. Hall 1,Stephen A. Moya 1,Marco A. Bauk 1,Jennifer J. Reeves 1,Mitchell C. Kong 1.11,Rany M. Salem 12,Gustavo Vizcardo-Galindo 3,Jose-Luis-Luis Macar Lupu 3,Romulo Figuero fox groude 3. Ikko Salomaa 14,Aki S. Havulinna 14.15,Andrew J. Murray 6,Atul Malhotra 1,Frank L. Powel 1,Mohit Jain 0,Alexis C. Komor 7,Gianpiero L.Cavalleri 4,Chad D.

EPAS1基因适应性遗传变异与世居高原藏族脑结构和 ...
土著藏族已经开发了自适应生理机制,以应对Qinghai-Xizang高原的低氧环境。据报道,与缺氧诱导因子途径相关的内皮PAS蛋白1基因(EPAS1)内的遗传变异与藏族之间的低氧适应性有关。大脑在体内表现出最高的氧气消耗,特别容易受到高空缺氧的影响。我们研究了Qinghai-Xizang高原中藏族的结构和功能性脑网络的遗传影响。在这项研究中,招募了135名年轻土著藏族(62名男性和73名女性)作为实验组。 65名与相关特征相匹配的低地汉族人被招募为遗传变异分析的对照组。基于先前的报道,选择了EPAS1中的12个单核苷酸多态性基因座进行基因分型。随后,使用磁共振成像(MRI)获得了大脑的T1结构和静止状态功能图像。单倍型分析表明,藏族中GA和CAAA单倍型的频率明显高于低地汉族个体。藏人被认为是更高的适应性。因此,藏族被归类为遗传适应的藏族(GHA-tibetans)和遗传适应性较低的藏人(GLA-tibetans)。自适应的大脑变化也参与了自发的休息状态活动网络。与Gla-tibetans相比,Gha-tibetans在左中央回和右侧毛氨酸回去,右侧额叶和右后扣带回回去的皮质表面积明显更大,在左PericalCarine Gyrus和右PericalCarine Gyrus和右上角的皮质体积中,右侧额叶和右后扣回去。在多个网络中观察到功能连接显着提高,包括体育体网络,腹侧注意网络,视觉网络和默认模式网络。这项研究揭示了EPAS1遗传变异与土著藏族中大脑结构和功能网络的适应性之间的关系,表明大脑的适应性变化主要集中在与视觉感知,运动控制和相关功能网络相关的区域上。这些大脑变化可能有助于土著人口在极端环境中更好地调节其身体活动。
逆转癌症中的DNA高甲基化(综述)
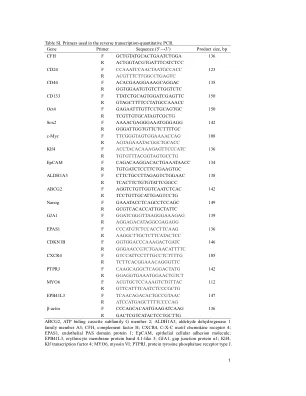
cfh f gctgtatgcactgaatctgga 136 r actgggtacgtgtgatttcatctccccccccccccccccccccccccccccccccccccccccccccccccc 123 r acgtttttttttcgctgcctgagtc cd44 f acacgagaagaagaagagagagcaggac 135 ttatctgcagtggatcgagttc 150 r gtagcttttcctttcctatgccaaacc oct4 f gagaatttgtgttgtcctggagtgc150 r tcgttgtgtgtgcatagtgctgtcgctgtcgcgtcggctg sox2 TTCGGGTAGTGGAAAACCAG 108 R AGTAGAAATACGGCTGCACC Klf4 F ACCTACACAAAGAGTTCCCATC 136 R TGTGTTTACGGTAGTGCCTG EpCAM F CAGACAAGGACACTGAAATAACC 134 R TGTGATCTCCTTCTGAAGTGC ALDH1A3 F cttctgccttagagtctggaac 138 r tcacttctgtgtgtattcggcc abcg2 f aggtctgtgtgtggtggtcaatctcac 142 r tcctgttgcattgagtcctg nanog nanog nanog f gaaatacctcctcctcagcctcctcctccctccagc149 ggatcgggttaagggaaagag 139 r aggagacataggcgagaggggggggggggg epas1 f cccatgtctccaccttcaag 136 r aaggcttgcttcttcattccttcatctcccccccccccccccccccacacaagcaagactc146 r gggggggggtccgtccccccctccctcctcccctcct4 105 r tcttcacggaaacagggttc ptprj f caagcaggctcaggactatg 142 r ggaggtgaAatggaAtggaActgtct myo6 f acgtgctccaaagtctgtgttac12 atccatgagcttttttccccagβ-肌动蛋白f cccagcacaatgaagatcaag 136 r gactcgtcatcatactcctgcttg abcg2,atp biding cassette cassette subfimily g ement g ement 2; Aldh1a3,醛脱氢酶1家族成员A3; CFH,补体因子H; CXCR4,C-X-C基序趋化因子受体4; EPAS1,内皮PAS结构域蛋白1; Epcam,上皮细胞粘附分子; EPB41L3,红细胞膜蛋白带4.1样3; GJA1,间隙连接蛋白α1; KLF4,KLF转录因子4; Myo6,肌球蛋白VI; PTPRJ,蛋白酪氨酸磷酸酶受体类型J

一种新的有效策略,是从人多能干细胞中产生肝窦内皮细胞
肝脏正弦内皮细胞(LSEC)是高度专业的内皮细胞(EC),在肝发育和再生中起着重要作用。此外,它参与了各种病理过程,包括脂肪变性,炎症,纤维化和肝细胞癌。然而,培养后LSEC的快速去分化极大地限制了其在生物医学应用中的体外建模。在这项研究中,我们开发了一种高效的方案,用于仅在8天内诱导人类诱导的多能干细胞(HIPSC)的LSEC像细胞。使用单细胞转录组分析,我们确定了几种新型LSEC特异性标记,例如EPAS1,LIFR和NID1,以及几种先前揭示的标记物,例如CLEC4M,CLEC1B,CRHBP,CRHBP和FCN3。这些LSEC标记在我们的LSEC样细胞中特异性表达。此外,HIPSC衍生的细胞表达LSEC特异性蛋白,并表现出与LSEC相关的功能,例如乙酰化低密度脂蛋白(AC-LDL)和免疫复杂的内吞作用。总体而言,这项研究证实了我们的新规程允许HIPSC迅速在体外获得LSEC样表型和功能。有效,迅速生成LSEC的能力可能有助于在肝特异性多细胞微环境中更精确地模仿肝发育和疾病进展,从而为新的治疗策略的发展提供新的见解。

缺氧诱导因子2α促进保护性TH2细胞...
Hypoxia-inducible factor 2 α promotes protective Th2 cell responses during intestinal 1 helminth infection 2 3 Jasmine C. Labuda 1 , Tayla M. Olsen 1,2 , Sheenam Verma 1 , Samantha Kimmel 1 , Thomas H. 4 Edwards 3 , Matthew J. Dufort 3 , Oliver J. Harrison 1,4 5 6 1 Center for Fundamental Immunology, Benaroya Research美国华盛顿州西雅图研究所。7 2分子和蜂窝生物学计划,美国华盛顿州西雅图市华盛顿大学。8 3美国华盛顿州西雅图市贝纳罗亚研究所系统免疫学中心。9 4美国华盛顿州华盛顿大学华盛顿大学免疫学系。10 11通信:oharrison@benaroyaresearch.org 12 13摘要:TH2细胞必须感知并适应组织环境,以提供保护性宿主14免疫和组织修复。在这里,我们检查了促进Th2细胞15分化和功能的机制。单细胞RNA-seq 16分析来自小肠道椎板椎板的CD4 + T细胞17揭示了基因EPAS1的高表达,编码了转录因子缺氧缺氧诱导的18因子2a(HIF2α)。在体外,即使在非极化条件下,暴露于缺氧或遗传HIF2α激活也促进了Th2细胞19分化。在小鼠中,CD4 + T细胞中的HIF2α激活20在没有感染的情况下促进了肠道Th2细胞的积累,而HIF2α缺陷21受损的CD4 + T细胞介导的宿主对肠舵感感染的免疫免疫。24 25简介:肠蠕虫感染是全球最普遍的慢性感染26。我们的发现22确定了缺氧,氧调节的转录因子缺氧诱导因子2α23(HIF2α)是小肠内Th2细胞分化和功能的关键调节剂。Helminth infections are often associated with polarized “type 2” immunity, including 27 activation and accumulation of T helper 2 (Th2) cells, type-2 innate lymphoid cells (ILC2), tissue 28 basophils and eosinophils, elevated serum immunoglobulin E (IgE), alternative activation of 29 macrophages and alterations of epithelial differentiation and mucus production that统称30重塑感染的解剖部位2。免疫事件和31个组织重塑的类似级联反应引发局部组织病理学发生在过敏性疾病中,包括过敏32哮喘3。33 34指导屏障组织中Th2细胞分化的机制尚不清楚。35然而,证据支持组织微环境在建立36保护性Th2细胞分化和功能中的指导性作用,这是由染色质访问性37和/或基因表达的变化提供的,在将Th2细胞从淋巴结到本塞质体38组织4,5的TH2细胞转运后的基因表达。组织警报蛋白,包括IL-25,IL-33和TSLP是在39个屏障组织中产生的关键因素,这些因素在Helminth 40感染6,7期间共同促进2型免疫力和Th2细胞反应。在41个屏障组织中影响Th2细胞功能的组织环境中其他提示的身份仍有待鉴定。42 43缺氧诱导因子(HIF)是介导细胞的关键转录因子,对缺氧8的有机体反应4。Consisting of 3 family members, (HIF1 α , HIF2 α and HIF3 α , 45 encoded by Hif1a , Epas1 and Hif3a, respectively), HIFs are post-translationally modified in an 46 oxygen-dependent enzymatic cascade that regulates their stability, nuclear translocation, 47 binding to hypoxia-response elements (HRE) and transcription of低氧诱导基因8。48在常氧条件下,HIF蛋白通过氧气在关键的脯氨酸残基上通过氧气-49依赖性丙酰羟化酶(PHD)酶羟基氧化。通过50

全基因组对中国土著绵羊的重新陈述提供了对遗传基础气候适应的洞察力
抽象背景中国土著绵羊是具有独特特征和特征的宝贵资源。它们分布在中国大陆的气候不同的地区;但是,很少有报道根据其基因组分析了绵羊的环境适应性。我们研究了适应于对极端湿度,高度和温度条件的选择的变体和特征,这些绵羊基因组中的41种表型和地理位置代表性的中国土著绵羊繁殖以表征这些种群中的遗传基础环境适应的遗传基础。基于人口结构分析的结果,我们推断中国土著绵羊分为四类:哈萨克(KAZ),蒙古人(MON),藏族(Tib)(Tib)和Yunnan(Yun)。我们还检测了一组与适应极端环境条件相关的罐头基因,例如易于干旱的区域(TBXT,TG,TG和HOXA1),高蓝色区域(DYSF,EPAS1,JAZF1,JAZF1,PDGFD,PDGFD和NF1和NF1和NF1)和温暖的区域(Tshr,tshr,abcD4)和ABCD4和ABCD4和ABCD4,在所有这些候选基因中,八个ABCD4,CNTN4,DOCK10,LOC105608545,LOC121816479,SEM3A,SVIL和TSHR在极端环境条件之间重叠。TSHR基因在温暖组中显示出强烈的签名,并在染色体上置于90,600,001和90,650,001之间的单个核苷酸聚合物(SNP)错义突变,这会导致TSHR蛋白质结构的变化,并影响其稳定性。对选择与环境适应性有关的选择基因和TSHR基因中SNP错义突变的结论分析,该基因影响蛋白质结构和稳定性。它还提供了有关中国土著绵羊种群植物地理结构演变的信息。这些结果为未来的繁殖研究提供了重要的遗传资源,以及有关动物如何适应气候变化的新观点。