XiaoMi-AI文件搜索系统
World File Search SystemHelicobacter

筛选小分子糖基化抑制剂并评估其对糖蛋白的影响
Bowdoin College生物化学系摘要:幽门螺杆菌是一种革兰氏阴性的,螺旋形的致病细菌,可在人类的胃中殖民。一旦建立了感染,还可能发生胃癌和消化性溃疡。抗生素一直是致病细菌治疗的强大工具。当前幽门螺杆菌的治疗方法是“三重治疗”,涉及至少两种不同的抗生素。第三种药物通常是质子泵抑制剂(PPI),以帮助胃愈合。但是,这尚未有效。过量使用广谱抗生素会导致细菌病原体中的抗生素耐药机制(Quintana,2022)。广谱抗生素也经常破坏正常的肠道微生物组(Quintana,2022)。需要替代方法来最大程度地减少负面副作用,并选择性地治疗关注的抗生素抗生素“优先病原体”。细菌聚糖由于其独特的单糖和在细菌致病性中的作用而引人注目的治疗靶标。先前的研究表明,细菌糖基化的破坏会导致对宿主细胞,生物膜形成,运动的粘附降低,从而降低了宿主定殖(Quintana,2022)。因此,杜贝实验室的研究重点是靶向覆盖细菌细胞表面的聚糖。伊莎贝拉·金塔纳(Isabella Quintana)最近的工作先前证明了基于稀有细菌单糖的S-糖苷抑制剂,可有效破坏聚糖生物合成和细菌适应性。我复制了一些实验以确认这些发现。我还使用碳水化合物结合蛋白探索了互补角,以确认S-糖苷抑制剂以浓度依赖性方式有效地阻碍了聚糖生物合成。总体而言,小分子抑制剂已证明可以选择性地靶向抗生素耐药性病原体,从而增加了我们治疗感染的工具。项目目标:这项研究旨在评估基于稀有细菌单糖的S-糖苷抑制剂在破坏聚糖生物合成中的有效性,通过测量糖蛋白生物合成和细胞表面聚糖结构的变化。使用的方法:小分子抑制剂 - 细菌单糖,BAC,DATDG和FUCNAC上的苄基 - 糖苷被用作细菌糖基转移酶的底物诱饵。这些细菌单糖是由于它们快速合成支架及其在优先病原体中的利用而选择的(Williams,2020)。目的是评估S-糖苷作为代谢抑制剂的有效性,以阻止细菌糖基化。这是通过1)确认抑制糖蛋白生物合成的抑制作用和2)确认细胞表面聚糖结构的变化。

马来西亚Tegoprazan的营销批准
2024年9月26日,日本马来西亚纳戈亚的Tegoprazan的营销批准 - 拉奎里亚(Raqualia Pharma Inc. Pharmaniaga Logistics Sdn Bhd的合作伙伴(总部:马来西亚Shah Alam,“ Pharmaniaga”)已获得国家药品监管机构(NPRA)的营销批准,胃酸分泌抑制剂Tegoprazan(“ Tegoprazan”)。tegoprazan是一种钾的抗酸性阻滞剂(P-CAB),一种新产生的酸抑制剂。p-cabs具有与质子泵抑制剂(PPI),胃食管反流疾病的一线疗法不同的作用机理,并且比PPI更快,更持久地抑制胃酸分泌。Raqualia和HK Inno.N签订了具有转态权利的开发,营销和制造Tegoprazan的独家许可协议。HK Inno.N及其全球业务合作伙伴一直在为Tegoprazan开展业务活动。目前,Tegoprazan产品在包括韩国,蒙古,中国,菲律宾,印度尼西亚,新加坡,墨西哥,秘鲁和智利的九个国家 /地区销售,并在包括美国在内的37个国家 /地区进行了发射,批准审查和临床试验的准备工作。在马来西亚,2021年,香港Inno.n与Pharmaniaga达成了成品供应协议,从那时起,Pharmaniaga一直在努力获得营销批准。批准的适应症包括侵蚀性胃食管反流疾病,非弱化性胃食管反流疾病,胃溃疡和幽门螺杆菌消除。当地市场上的品牌名称是K-Cab®,Pharmaniaga预计该产品将于2025年上半年推出。东南亚消化性溃疡药物的市场规模为5.2亿美元,预计将进一步增长。tegoprazan已经在菲律宾,新加坡和印度尼西亚以及现在获得批准的马来西亚销售,也在泰国和越南的监管审查过程中,在东南亚的经济规模上,在泰国和越南获得了批准的批准。根据与香港Inno.n的许可协议,拉奎里亚有权获得Sublicensees HK Inno.N的一部分收入。尽管由于此法规批准,拉奎里亚(Raqualia)将不收取一次性付款,但拉奎里亚(Raqualia)认为,扩大Tegoprazan销售的国家数量将有助于在长期到长期中提高我们的业务收入和公司价值。Raqualia将继续加强与HK Inno.N的合作伙伴关系,为开发和偶然成分提供持续的支持,以扩大与胃酸相关疾病的选择,从而为改善患者的生活质量做出了进一步的贡献。

细菌植物的优点和缺点
4。Ansaldo E,Slayden LC,Ching KL,Koch MA,Wolf NK,Plichta DR等。akkermansia粘膜粘膜在稳态期间诱导肠道适应性免疫反应。科学。2019; 364(6446):1179-1184。 5。 Sefik E,Geva-Zatorsky N,Oh S,Konnikova L,Zemmour D,McGuire AM等。 个体肠道共生体诱导RORγ +调节性T细胞的不同种群。 科学。 2015; 349(6251):993-997。 6。 Lathrop SK,Bloom SM,Rao SM,Nutsch K,Lio CW,Santacruz N等。 结肠共生微生物群对免疫系统的外围教育。 自然。 2011; 478(7368):250-254。 7。 Yang Y,Torchinsky MB,Gobert M,Xiong H,Xu M,Linehan JL等。 肠道Th17细胞对共生细菌抗原的聚焦特异性。 自然。 2014; 510(7503):152-156。 8。 Xu M,Pokrovskii M,Ding Y,Yi R,Au C,Harrison OJ等。 c- MAF依赖性调节性T细胞介导对肠道病原体的免疫耐受性。 自然。 2018; 554(7692):373-377。 9。 Chai JN,Peng Y,Rengarajan S,Solomon BD,AI TL,Shen Z等。 螺旋杆菌是体内平衡和炎症中结肠T细胞反应的有效驱动因素。 SCI免疫。 2017; 2(13):EAAL5068。 10。 Ivanov II,Atarashi K,Manel N,Brodie EL,Shima T,Karaoz U等。 通过分段丝状细菌诱导肠道Th17细胞。 单元格。 2009; 139(3):485-498。 11。2019; 364(6446):1179-1184。5。Sefik E,Geva-Zatorsky N,Oh S,Konnikova L,Zemmour D,McGuire AM等。个体肠道共生体诱导RORγ +调节性T细胞的不同种群。科学。2015; 349(6251):993-997。 6。 Lathrop SK,Bloom SM,Rao SM,Nutsch K,Lio CW,Santacruz N等。 结肠共生微生物群对免疫系统的外围教育。 自然。 2011; 478(7368):250-254。 7。 Yang Y,Torchinsky MB,Gobert M,Xiong H,Xu M,Linehan JL等。 肠道Th17细胞对共生细菌抗原的聚焦特异性。 自然。 2014; 510(7503):152-156。 8。 Xu M,Pokrovskii M,Ding Y,Yi R,Au C,Harrison OJ等。 c- MAF依赖性调节性T细胞介导对肠道病原体的免疫耐受性。 自然。 2018; 554(7692):373-377。 9。 Chai JN,Peng Y,Rengarajan S,Solomon BD,AI TL,Shen Z等。 螺旋杆菌是体内平衡和炎症中结肠T细胞反应的有效驱动因素。 SCI免疫。 2017; 2(13):EAAL5068。 10。 Ivanov II,Atarashi K,Manel N,Brodie EL,Shima T,Karaoz U等。 通过分段丝状细菌诱导肠道Th17细胞。 单元格。 2009; 139(3):485-498。 11。2015; 349(6251):993-997。6。Lathrop SK,Bloom SM,Rao SM,Nutsch K,Lio CW,Santacruz N等。结肠共生微生物群对免疫系统的外围教育。自然。2011; 478(7368):250-254。7。Yang Y,Torchinsky MB,Gobert M,Xiong H,Xu M,Linehan JL等。 肠道Th17细胞对共生细菌抗原的聚焦特异性。 自然。 2014; 510(7503):152-156。 8。 Xu M,Pokrovskii M,Ding Y,Yi R,Au C,Harrison OJ等。 c- MAF依赖性调节性T细胞介导对肠道病原体的免疫耐受性。 自然。 2018; 554(7692):373-377。 9。 Chai JN,Peng Y,Rengarajan S,Solomon BD,AI TL,Shen Z等。 螺旋杆菌是体内平衡和炎症中结肠T细胞反应的有效驱动因素。 SCI免疫。 2017; 2(13):EAAL5068。 10。 Ivanov II,Atarashi K,Manel N,Brodie EL,Shima T,Karaoz U等。 通过分段丝状细菌诱导肠道Th17细胞。 单元格。 2009; 139(3):485-498。 11。Yang Y,Torchinsky MB,Gobert M,Xiong H,Xu M,Linehan JL等。肠道Th17细胞对共生细菌抗原的聚焦特异性。自然。2014; 510(7503):152-156。 8。 Xu M,Pokrovskii M,Ding Y,Yi R,Au C,Harrison OJ等。 c- MAF依赖性调节性T细胞介导对肠道病原体的免疫耐受性。 自然。 2018; 554(7692):373-377。 9。 Chai JN,Peng Y,Rengarajan S,Solomon BD,AI TL,Shen Z等。 螺旋杆菌是体内平衡和炎症中结肠T细胞反应的有效驱动因素。 SCI免疫。 2017; 2(13):EAAL5068。 10。 Ivanov II,Atarashi K,Manel N,Brodie EL,Shima T,Karaoz U等。 通过分段丝状细菌诱导肠道Th17细胞。 单元格。 2009; 139(3):485-498。 11。2014; 510(7503):152-156。8。Xu M,Pokrovskii M,Ding Y,Yi R,Au C,Harrison OJ等。c- MAF依赖性调节性T细胞介导对肠道病原体的免疫耐受性。自然。2018; 554(7692):373-377。 9。 Chai JN,Peng Y,Rengarajan S,Solomon BD,AI TL,Shen Z等。 螺旋杆菌是体内平衡和炎症中结肠T细胞反应的有效驱动因素。 SCI免疫。 2017; 2(13):EAAL5068。 10。 Ivanov II,Atarashi K,Manel N,Brodie EL,Shima T,Karaoz U等。 通过分段丝状细菌诱导肠道Th17细胞。 单元格。 2009; 139(3):485-498。 11。2018; 554(7692):373-377。9。Chai JN,Peng Y,Rengarajan S,Solomon BD,AI TL,Shen Z等。螺旋杆菌是体内平衡和炎症中结肠T细胞反应的有效驱动因素。SCI免疫。 2017; 2(13):EAAL5068。 10。 Ivanov II,Atarashi K,Manel N,Brodie EL,Shima T,Karaoz U等。 通过分段丝状细菌诱导肠道Th17细胞。 单元格。 2009; 139(3):485-498。 11。SCI免疫。2017; 2(13):EAAL5068。 10。 Ivanov II,Atarashi K,Manel N,Brodie EL,Shima T,Karaoz U等。 通过分段丝状细菌诱导肠道Th17细胞。 单元格。 2009; 139(3):485-498。 11。2017; 2(13):EAAL5068。10。Ivanov II,Atarashi K,Manel N,Brodie EL,Shima T,Karaoz U等。 通过分段丝状细菌诱导肠道Th17细胞。 单元格。 2009; 139(3):485-498。 11。Ivanov II,Atarashi K,Manel N,Brodie EL,Shima T,Karaoz U等。通过分段丝状细菌诱导肠道Th17细胞。单元格。2009; 139(3):485-498。 11。2009; 139(3):485-498。11。Bilate AM,Bousbaine D,Mesin L,Agudelo M,Leube J,Kratzert A等。来自克隆T细胞前体的调节和上皮内T细胞的组织特异性出现。SCI免疫。 2016; 1(2):EAAF7471。 12。 Bilate Am,Lafaille JJ。 在免疫耐受性中诱导的CD4+ FOXP3+调节T细胞。 Annu Rev Immunol。 2012; 30:733-758。 13。 页岩M,Schiering C,Powrie F.肠炎中的CD 4+ T细胞子集。 Immunol Rev. 2013; 252(1):164-182。 14。 Sujino T,伦敦M,Hoytema van Konijnenburg DP,Rendon T,Buch T,Silva HM等。 调节和上皮内CD4+ T细胞的组织适应控制肠道炎症。 科学。 2016; 352(6293):1581-1586。 15。 Reis BS,Rogoz A,Costa-Pinto FA,Taniuchi I,MucidaD。转录因子Runx3和ThPOK的相互表达调节肠道CD4+ T细胞免疫。 nat免疫。 2013; 14(3):271-280。SCI免疫。2016; 1(2):EAAF7471。 12。 Bilate Am,Lafaille JJ。 在免疫耐受性中诱导的CD4+ FOXP3+调节T细胞。 Annu Rev Immunol。 2012; 30:733-758。 13。 页岩M,Schiering C,Powrie F.肠炎中的CD 4+ T细胞子集。 Immunol Rev. 2013; 252(1):164-182。 14。 Sujino T,伦敦M,Hoytema van Konijnenburg DP,Rendon T,Buch T,Silva HM等。 调节和上皮内CD4+ T细胞的组织适应控制肠道炎症。 科学。 2016; 352(6293):1581-1586。 15。 Reis BS,Rogoz A,Costa-Pinto FA,Taniuchi I,MucidaD。转录因子Runx3和ThPOK的相互表达调节肠道CD4+ T细胞免疫。 nat免疫。 2013; 14(3):271-280。2016; 1(2):EAAF7471。12。Bilate Am,Lafaille JJ。在免疫耐受性中诱导的CD4+ FOXP3+调节T细胞。Annu Rev Immunol。 2012; 30:733-758。 13。 页岩M,Schiering C,Powrie F.肠炎中的CD 4+ T细胞子集。 Immunol Rev. 2013; 252(1):164-182。 14。 Sujino T,伦敦M,Hoytema van Konijnenburg DP,Rendon T,Buch T,Silva HM等。 调节和上皮内CD4+ T细胞的组织适应控制肠道炎症。 科学。 2016; 352(6293):1581-1586。 15。 Reis BS,Rogoz A,Costa-Pinto FA,Taniuchi I,MucidaD。转录因子Runx3和ThPOK的相互表达调节肠道CD4+ T细胞免疫。 nat免疫。 2013; 14(3):271-280。Annu Rev Immunol。2012; 30:733-758。 13。 页岩M,Schiering C,Powrie F.肠炎中的CD 4+ T细胞子集。 Immunol Rev. 2013; 252(1):164-182。 14。 Sujino T,伦敦M,Hoytema van Konijnenburg DP,Rendon T,Buch T,Silva HM等。 调节和上皮内CD4+ T细胞的组织适应控制肠道炎症。 科学。 2016; 352(6293):1581-1586。 15。 Reis BS,Rogoz A,Costa-Pinto FA,Taniuchi I,MucidaD。转录因子Runx3和ThPOK的相互表达调节肠道CD4+ T细胞免疫。 nat免疫。 2013; 14(3):271-280。2012; 30:733-758。13。页岩M,Schiering C,Powrie F.肠炎中的CD 4+ T细胞子集。Immunol Rev.2013; 252(1):164-182。 14。 Sujino T,伦敦M,Hoytema van Konijnenburg DP,Rendon T,Buch T,Silva HM等。 调节和上皮内CD4+ T细胞的组织适应控制肠道炎症。 科学。 2016; 352(6293):1581-1586。 15。 Reis BS,Rogoz A,Costa-Pinto FA,Taniuchi I,MucidaD。转录因子Runx3和ThPOK的相互表达调节肠道CD4+ T细胞免疫。 nat免疫。 2013; 14(3):271-280。2013; 252(1):164-182。14。Sujino T,伦敦M,Hoytema van Konijnenburg DP,Rendon T,Buch T,Silva HM等。调节和上皮内CD4+ T细胞的组织适应控制肠道炎症。科学。2016; 352(6293):1581-1586。 15。 Reis BS,Rogoz A,Costa-Pinto FA,Taniuchi I,MucidaD。转录因子Runx3和ThPOK的相互表达调节肠道CD4+ T细胞免疫。 nat免疫。 2013; 14(3):271-280。2016; 352(6293):1581-1586。15。Reis BS,Rogoz A,Costa-Pinto FA,Taniuchi I,MucidaD。转录因子Runx3和ThPOK的相互表达调节肠道CD4+ T细胞免疫。nat免疫。2013; 14(3):271-280。2013; 14(3):271-280。

质子泵抑制剂与癌症:现状
幽门螺杆菌感染和 Zollinger-Ellison 综合征,以及预防高危患者(年龄 > 65 岁,有胃肠道溃疡病史或同时接受抗血小板、抗凝或皮质类固醇治疗)的非甾体抗炎药 (NSAID) 相关胃肠道病变(Strand 等人,2017 年)。许多此类疾病通常需要长期治疗,这增加了患者发生临床上显著的药物相互作用的可能性。此外,标签外处方已被广泛报道,特别是在功能性消化不良和预防非风险患者 NSAID 引起的胃十二指肠病变方面(Lassalle 等人,2020 年)。自 1980 年代后期推出市场以来,PPI 在许多国家的使用都有所增加。例如,在法国,2015 年有超过 1500 万拥有医疗保险的人(占法国成年人口的近三分之一)是 PPI 使用者(Singh 等人,2018 年;Lassalle 等人,2020 年)。在一项研究中,三分之一的患者无法确定 PPI 指征,四分之三的 NSAID 预防性处方没有发现可测量的风险因素(Lassalle 等人,2020 年)。大约 20% 的癌症患者使用 PPI(Kinoshita 等人,2018 年;Tvingsholm 等人,2018 年;Sharma 等人,2019 年);然而,PPI 通常会被过量用于治疗化疗的副作用(如 GERD)或作为与皮质类固醇或 NSAID 联合治疗的预防措施(Lassalle 等人,2020 年)。总体而言,PPI 被认为不良事件很少,因为它们通常耐受性良好。然而,据报道,PPI 与胃肠道疾病(恶心、腹痛、传输障碍)、离子吸收障碍(低镁血症、铁缺乏、维生素 B12 缺乏)、肾衰竭、感染(肺炎、艰难梭菌感染、腹膜炎)和骨折有关(Singh 等人,2018 年;Yibirin 等人,2021 年)。此外,PPI 还参与各种药物 - 药物相互作用 (DDI) (Wedemeyer 和 Blume,2014 年;Strand 等人,2017 年;Patel 等人,2020 年;Uchiyama 等人,2021 年)。通过提高胃液 pH 值,PPI 会影响胃液 pH 依赖性药物的吸收。事实上,某些弱碱性药物的胃液 pH 值升高会导致溶解度降低,随后的吸收率也会降低 (Wedemeyer 和 Blume,2014 年;Patel 等人,2020 年)。PPI 也可能影响药物消除,因为它们是有机阳离子转运蛋白 (OCT,参与底物药物的肾脏排泄) 和 P-糖蛋白流出转运蛋白的潜在抑制剂 (Wedemeyer 和 Blume,2014 年;Patel 等人,2020 年)。 PPI 主要在肝脏中通过细胞色素 P450 酶 (CYP) 系统代谢,主要是 CYP2C19 和 CYP3A4 ( Wedemeyer and Blume, 2014 )。它们能够作为 CYP 的抑制剂或诱导剂;抑制 CYP 会增加全身对药物的暴露量 (Patel 等人,2020 年)。奥美拉唑对 CYP2C19 具有高亲和力,对 CYP3A4 具有中等亲和力,因此具有相当大的 DDI 潜力 (Wedemeyer and Blume, 2014 年)。埃索美拉唑也能在临床上显著抑制 CYP2C19,而其他 PPI 对 CYP2C19 的抑制在临床上并不重要 (Patel 等人,2020 年)。然而,只有少数涉及 PPI 的 DDI 具有临床意义 (Wedemeyer and Blume, 2014 年)。尽管如此,
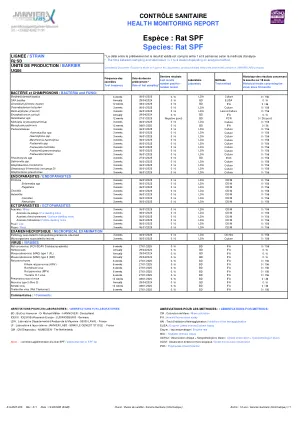
大鼠SPF
BACTERIA and FUNGI / BACTERIA and FUNGI Bordetella bronchiseptica 3 weeks 06/01/2025 0/6 LDA Culture 0/156 CAR bacillus Annually 29/04/2024 0/6 BD ELISA 0/6 Clostridium piliforme (tyzzer) 12 weeks 5 0/6 BD IFA 0/36 Corynebacterium kutscheri 3 weeks 06/01/2025 0/6 LDA Culture 0/156 Dermatophytes (if lesion) 3 weeks 0/01/2025 0/6 LDA Lesion/Culture 0/156 Encephalitozoon cuniculi 29/04/2025 Annually. 2024 0/6 BD IFA 0/6 Helicobacter spp 12 weeks 27/01/2025 Negative (pool) BD PCR 0/26 (pool) Klebsiella oxytoca/pneumoniae 3 weeks 0/01/2025 0/6 LDA Culture 0/156 Mycoplasma pulmonis 12 weeks. 01/2025 0/6 BD IFA 0/36 Pasteurellaceae 3 weeks 06/01/2025 0/6 LDA Culture 0/156 Actinobacillus spp. 3 周 2025 年 6 月 1 日 0/6 LDA 培养 0/156 嗜血杆菌属。 3 周 06/01/2025 0/6 LDA 培养 0/156 溶血曼海姆氏菌 3 周 06/01/2025 0/6 LDA 培养 0/156 巴氏杆菌属。 3 周 06/01/2025 0/6 LDA 培养 0/156 多杀性巴氏杆菌 3 周 0/01/2025 0/6 LDA 培养 0/156 嗜肺巴氏杆菌 3 周 0/01/2025 0/6 LDA 培养 0/156 海藻巴氏杆菌 周 06/01/2025 0/6 LDA 培养 0/156 肺孢子菌属。 3 周 2025 年 1 月 27 日 0/6 BD PCR 0/156 沙门氏菌属。3 周 06/01/2025 0 / 6 LDA 培养 0 / 156 念珠状链杆菌 3 周 06/01/2025 0 / 6 LDA 培养 0 / 156 β-溶血性链球菌(非 D 组) 3 周 06/01/2025 0 / 6 LDA 培养 0 / 156 肺炎链球菌 3 周 06/01/2025 0 / 6 LDA 培养 0 / 156 体内寄生虫 / 体内寄生虫 原生动物 3 周 06/01/2025 0 / 6 LDA OD/M 0 / 156 内阿米巴属 3 周 06/01/2025 0 / 6 LDA OD/M 0 / 156 鞭毛虫3 周 06/01/2025 0 / 6 LDA OD/M 0 / 156 球虫 3 周 06/01/2025 0 / 6 LDA OD/M 0 / 156 蠕虫 3 周 06/01/2025 0 / 6 LDA OD/M 0 / 156 绦虫 3 周 06/01/2025 0 / 6 LDA OD/M 0 / 156 线虫 3 周 06/01/2025 0 / 6 LDA OD/M 0 / 156 体外寄生虫 / 体外寄生虫 螨虫 3 周 06/01/2025 0 / 6 LDA OD/M 0 / 156 皮螨 / 毛螨 3 周06/01/2025 0 / 6 LDA OD/M 0 / 156 环境螨 / 表面螨虫 3 周 06/01/2025 0 / 6 LDA OD/M 0 / 156 毛囊螨 / 毛囊螨 3 周 06/01/2025 0 / 6 LDA OD/M 0 / 156 虱子/虱子 3 周 06/01/2025 0 / 6 LDA OD/M 0 / 156 虱子/跳蚤 3 周 06/01/2025 0 / 6 LDA OD/M 0 / 156 检查 尸检/尸检 与观察到的组织病理学病变相关的病理学3周2025 年 1 月 6 日 0 / 6 LDA Ob/Hist 0 / 156 与病变相关的微生物 3 周 2025 年 1 月 6 日 0 / 6 LDA 培养 0 / 156 病毒 大鼠冠状病毒 (RCV/SDAV 涎腺腺炎) 6 周 2025 年 1 月 27 日 0 / 6 BD IFA 0 / 156 汉坦病毒 每年 2024 年 4 月 29 日 0 / 6 BD IFA 0 / 6 小鼠腺病毒 (MAD) 1 型 (FL) 每年 2024 年 4 月 29 日 0 / 6 BD IFA 0 / 6 小鼠腺病毒 (MAD) 2 型 (K87) 每年 2024 年 4 月 29 日 0 / 6 BD IFA 0 / 6 大鼠细小病毒 6 周2025/01/27 0 / 6 BD IFA 0 / 156 Kilham 大鼠细小病毒 (KRV) 6 周 2025/01/27 0 / 6 BD IFA 0 / 156 大鼠细小病毒 6 周 2025/01/27 0 / 6 BD IFA 0 / 156 大鼠细小病毒 (RPV) 6 周 2025/01/27 0 / 6 BD IFA 0 / 156 Toolan 的 H-1 病毒 6 周 2025/01/27 0 / 6 BD IFA 0 / 156 小鼠肺炎病毒 12 周 2025/01/08 0 / 6 BD IFA 0 / 36 呼肠孤病毒 3 型 (Reo 3) 每年 2024/04/29 0 / 6 BD IFA 0 / 6 仙台病毒 12 周 08/01/2025 0 / 6 BD IFA 0 / 36 类泰勒病毒 ('大鼠泰勒病毒') 6 周 27/01/2025 0 / 6 BD IFA 0 / 156

智力天赋和典型发育的记忆系统的神经解剖学差异
Huang-Pollock, CL、Maddox, WT 和 Karalunas, SL (2011)。内隐和外显类别学习的发展。《实验儿童心理学杂志》,109,321–335。 Kalbfleisch, ML (2004)。天赋的功能神经解剖学。《解剖记录》B 部分,277,21–36。 Kuhn, T.、Schonfeld, D.、Sayegh, P.、Arentoft, A.、Jones, JD、Hinkin, CH、Bookheimer, SY 和 Thames, AD (2017)。艾滋病毒和衰老对皮层下形状改变的影响:一项 3D 形态学研究。《人脑映射》,38(2),1025–1037。 Kyllonen, PC 和 Christal, RE (1990)。推理能力(仅仅)是工作记忆能力?智力,14,389–433。Laugeson, EA、Frankel, F.、Gantman, A.、Dillon, AR 和 Mogil, C. (2012)。针对患有自闭症谱系障碍的青少年的循证社交技能培训:UCLA PEERS 计划。自闭症和发育障碍杂志,42 (6),1025–1036。Mills, CJ 和 Tissot, SL (1995)。识别来自弱势群体学生的学术潜力:使用瑞文斯渐进矩阵是个好主意吗?天才儿童季刊,39,209–217。 Na, HS, Hong, SJ, Yoon, HJ, Maeng, JH, Ko, BM, Jung, IS, Ryu, CB, Kim, JO, Cho, JY, Lee, JS, Lee, MS, Shim, CS, & Kim, BS (2007)。幽门螺杆菌感染一线和二线治疗的根除率以及成功根除后的再感染率。韩国胃肠病学杂志,50,170-175。Navas-Sánchez, FJ, Alemán-Gómez, Y., Sánchez-Gonzalez, J., Guzmán-De-Villoria, JA, Franco, C., Robles, O., Arango, C., & Desco, M. (2014)。白质微结构与数学天赋和智商的关系。人脑映射,35(6),2619-2631。Neihart, M.、Reis, SM、Robinson, N. 和 Moon, S. (2002)。天才儿童的社交和情感发展:我们知道什么?Sourcebooks, Inc.O'Boyle,MW(2008)。数学天才儿童:大脑发育特征及其幸福感预后。Roeper Review,30(3),181-186。O'Boyle, MW、Alexander, JE 和 Benbow, CP (1991)。数学早熟儿童右半球活动增强:初步脑电图调查。脑与认知,17,138-153。O'Boyle, MW 和 Benbow, CP (1990)。认知处理过程中右半球参与度的提高可能与智力早熟有关。《神经心理学》,28,211-216。O'Boyle, MW、Benbow, CP 和 Alexander, JE (1995)。智力超常者的性别差异、半球侧化和相关大脑活动。《发育神经心理学》,11 (4),415-443。O'Boyle, MW、Cunnington, R.、Silk, TJ、Vaughan, D.、Jackson, G.、Syngeniotis, A. 和 Egan, GF (2005)。数学天才的男性青少年在心理旋转过程中激活独特的大脑网络。《认知脑研究》,25 (2),583-587。Packard, MG 和 Knowlton, BJ (2002)。基底神经节的学习和记忆功能。《神经科学年度评论》,25,563–593。
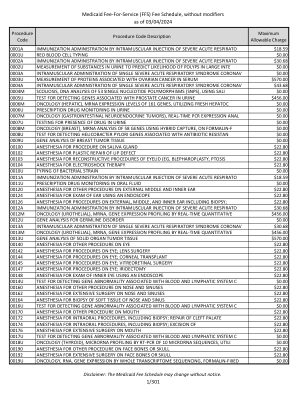
医疗补助按服务收费 (FFS) 费用表,不含修饰语
0001A 肌肉注射严重急性呼吸道综合征冠状病毒 (SARS) 进行免疫接种 $18.59 0001U 红细胞分型 $0.00 0002A 肌肉注射严重急性呼吸道综合征冠状病毒 (SARS) 进行免疫接种 $30.68 0002U 测量尿液中的物质以预测大肠息肉的可能性 $0.00 0003A 肌肉注射单个严重急性呼吸道综合征冠状病毒 (SARS) $0.00 0003U 测量血清中与卵巢癌相关的蛋白质 $570.00 0004A 肌肉注射单一严重急性呼吸道综合征冠状病毒的管理 $43.68 0004M 脊柱侧弯,使用 SALI 对 53 个单核苷酸多态性 (SNPS) 进行 DNA 分析 $0.00 0005U 测试用于检测尿液中与前列腺癌相关的基因 $456.00 0006M 肿瘤学 (肝脏),利用新鲜肝细胞检测 161 个基因的 mRNA 表达水平 $0.00 0006U 尿液中的处方药监测 $0.00 0007M 肿瘤学 (胃肠道神经内分泌肿瘤),实时 PCR 表达肛门 $0.00 0007U 检测尿液中是否存在药物 $0.00 0008M 肿瘤学(乳腺),使用杂交捕获法对福尔马林-F 上的 58 个基因进行 mRNA 分析 $0.00 0008U 检测与抗生素耐药性相关的幽门螺杆菌基因 $0.00 0009U 乳腺肿瘤组织的基因分析 $0.00 00100 唾液腺手术麻醉 $22.80 00102 唇缺损整形修复麻醉 $22.80 00103 眼睑重建手术麻醉(例如,眼睑成形术、眼睑下垂 $22.80 00104 电击疗法麻醉 $22.80 0010U 细菌菌株分型 $0.00 0011A 严重急性呼吸道感染肌肉注射免疫 $18.59 0011U 口服液处方药监测 $0.00 00120 外耳、中耳和内耳其他手术麻醉 $22.80 00124 使用内窥镜检查耳朵的麻醉 $22.80 00126 外耳、中耳和内耳手术麻醉包括活检; $22.80 0012A 通过肌肉注射严重急性呼吸道综合征冠状病毒进行免疫接种 $30.68 0012M 肿瘤学(尿路上皮),mRNA,通过实时定量基因表达分析 $456.00 0012U 种系疾病基因分析 $0.00 0013A 肌肉注射单一严重急性呼吸道综合征冠状病毒 $30.68 0013M 肿瘤学(尿路上皮),mRNA,通过实时定量基因表达分析 $456.00 0013U 实体器官肿瘤组织的基因分析$570.00 00140 其他眼部手术麻醉 $22.80 00142 眼部手术麻醉;晶状体手术 $22.80 00144 眼部手术麻醉;角膜移植 $22.80 00145 眼部手术麻醉;玻璃体视网膜手术 $22.80 00147 眼部手术麻醉;虹膜切除术 $22.80 00148 使用内窥镜检查眼内麻醉 $22.80 0014U 用于检测与血液和淋巴系统相关的基因异常的测试 C $0.00 00160 对鼻子和鼻窦的其他程序的麻醉 $22。80 00162 鼻腔和鼻窦大范围手术麻醉 $22.80 00164 鼻腔和鼻窦软组织活检麻醉 $22.80 0016U 血液和淋巴系统相关基因异常检测测试 C $0.00 00170 其他口腔手术麻醉 $22.80 00172 口腔内手术麻醉,包括活检;腭裂修复 $22.80 00174 口腔内手术麻醉,包括活检;切除术 $22.80 00176 口腔大范围手术麻醉 $22.80 0017U 血液和淋巴系统 C 相关基因异常检测测试 $0.00 0018U 肿瘤学(甲状腺),通过 RT-PCR 对 10 个微小 RNA 序列进行微小 RNA 分析,实用 $0.00 00190 面骨或头骨其他手术麻醉 $22.80 00192 面骨或头骨大范围手术麻醉 $22.80 0019U 肿瘤学,RNA,通过全转录组测序进行基因表达,福尔马林固定 $0.00
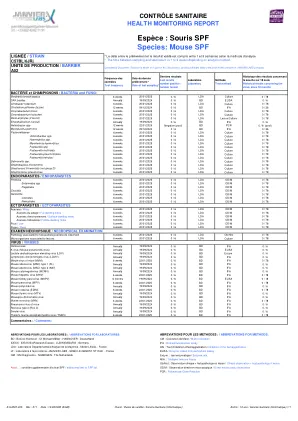
物种:SPF鼠标
Bactérieet Champignons /细菌和Fung Bonchiseptica 6周20/01/2025 0 /6 LDA文化Rodentium 6周20/01/2025 0/6 LDA文化0/78梭状芽胞杆菌(Tyzzer)12周09/12/2024 0/6 BD IFA 0/36 BOVIS BOVIS BOVIS 6周20/01/2025 0/2025 0/2025 0/6 LDA培养0 /78 Cownebacterium Kutscheri 6周20/01/2025 0 /6 LDA培养0/78皮肤植物(如果病变)年度19/09/2024 0/6 BD灯0/6 Helicabacter SPP 12周22/01/2025负(POLOL)BD 0/6(POL)Mycoplasma plemonis 12 Weeks 09/12/2024 0/6 BD IFAA 0/6 BD IFAA 0/ 36巴氏菌科6周20/01/2025 0 /6 LDA培养0/78 actinobcillus spp。6周20/01/2025 0 /6 LDA文化0/78 Heamophilus spp。6周20/01/2025 0 /6 LDA培养0/78 Mannheimia Haemytica Haemytica 6周20/01/2025 0 /6 LDA培养0/78 Pasteurella spp。6周20/01/2025 0/6 LDA培养0/78糊剂Multocida 6周20/01/2025 0/6 LDA培养0/78 pneumotropica pereumotropica 6 Weeks 20/01/2025 0/6 LDA LDA培养0 /78 treehalosi的巴斯德奶酪6周20/01/2025 0 /6 LDA培养0/78 Salmonnella spp。6 weeks 20/01/2025 0 / 6 LDA Culture 0 / 78 Streptobacillus moniliformis 6 weeks 20/01/2025 0 / 6 LDA Culture 0 / 78 Streptococci ß-hemolytic (not group D) 6 weeks 20/01/2025 0 / 6 LDA Culture 0 / 78 Streptococcus pneumoniae 6 weeks 20/01/2025 0 / 6 LDA Culture 0 / 78 ENDOPARASITES / ENDOPARASITES Protozoa 6 weeks 20/01/2025 0 / 6 LDA OD/M 0 / 78 Entamoeba spp 6 weeks 20/01/2025 0 / 6 LDA OD/M 0 / 78 Flagellates 6 weeks 20/01/2025 0 / 6 LDA OD/M 0 / 78 Coccidia 6 weeks 20/01/2025 0 / 6 LDA OD/M 0/78蠕虫6周20/01/2025 0/6 LDA OD/M 0/78 CESTODES 6周6周20/01/2025 0/6 LDA OD/M 0/M 0/M 0/M 0/78 NEMATODES 6 WEDWEN 20/01/01/01/2025 0/6 LDA OD/M 0/M 0/M 0/M 0/78 MIMESITES 6周6周6周6周6周20/01/2025 0/6 LDA OD/ M 0/78 Acariens du pelage/ tur-dewelling螨虫6周20/01/2025 0/6 LDA OD/ M 0/ M 0/ M 0/78 Acariens d'Acariens d'evironnement/ Surface-D'Erviration/ Surface-Deblist-Dewelling Mecites 6 Weeking 6 Weech weeks 20/01/2025 0 / 6 LDA OD/M 0 / 78 Poux / Lice 6 weeks 20/01/2025 0 / 6 LDA OD/M 0 / 78 Puces / Fleas 6 weeks 20/01/2025 0 / 6 LDA OD/M 0 / 78 EXAMEN NÉCROPSIQUE / NECROPSICAL EXAMINATION Pathology associated to histopathological lesions observed 6 weeks 20/01/2025 0 / 6 LDA OB / HIST 0 /78与病变相关的微生物6周20/01/2025 0 /6 LDA培养0/78每年19/09/2024 0 /6 BD IFA 0 /6 K病毒(小鼠肺炎病毒)每年19/6 k病毒hantaviruess hantaviruses每年19/09/09/09/20/20/2024 0 /6 BD D.病毒(LDV)每年19/09/2024 0/6 BD酶。0 /6每年19/09/2024每年19/09/0 / 6 ifa IFA 0/6分钟病毒(MVM)6周22/01/2025 0 /6 BD IFA IFA 0 /78小鼠adenovirus(MAD)IFA(MAD)IFA IFA IFA IFA EDA IFA IFA 0 /6 BDA 0 /6 BDA 0 /6 BDA 0 /6 BD ANEN / 6 BDIRIUL (MAD)类型2(K87)每年19/09/2024 0 /6 bd IFA 0/6鼠标cytomegalovirus(MCMV)每年19/09/2024 0 /6 BD IFA 0 /6 IFA 0 /6鼠标肝炎病毒病毒(MHV)6周6周22/01/2025 0 /6 BD IFA 0 /78 IFA 0 /78 MOUSS NEDNEY(MOK) 19/09/2024 0 /6 BD ELISA 0/18鼠标细小病毒(MPV)6周6周22/01/2025 0 /6 BD IFA 0/78鼠标多组病每年19/09/2024 0 /6 BD IFA 0/6 ifa 0 /6 Mouse Rotavirus(Edim)6周22/01/201/201/2025 0 /6 BD Annually 19/09/2024 0 / 6 BD IFA 0 / 6 Mousepox (Ectromelia) virus Annually 19/09/2024 0 / 6 BD IFA 0 / 6 Murine norovirus (MNV) 6 weeks 22/01/2025 0 / 6 BD IFA 0 / 78 Pneumonia virus of mice Annually 19/09/2024 0 / 6 BD IFA 0 / 6 Reovirus 3型(REO 3)每年19/09/2024 0/6 BD IFA 0/6每年19/09/2024 0/6 BD IFA 0/6 Theiler的鼠脑瘤瘤病毒病毒(TMEV)6周6周22/01/2025 0/6 BD IFA 0/6 BD IFA 0/78
![b'[2] C. Yan,X。Duanmu,L。Zeng,B。Liu,Z。Song,线粒体DNA:分布,突变和消除,细胞,8(2019)。 [3] F. Liu,D.E。 Sanin,X。Wang,肺癌中的线粒体DNA,实验医学和生物学进展,1038(2017)9-22。 [4] J. Zhang,J。 Jia,M。Junaid,Y.B。 MA,F。Ahmad,Y.F。 Jia,W.G。Li,D.S。Pei,人类DNA2(HDNA2)作为癌症和其他疾病的潜在靶标的作用:系统评价,DNA修复(AMST),59(2017)9-19。 [6] A. D \ XC3 \ Xadaz-Talavera,C。Montero-Conde,L.J。Leandro-Garc \ XC3 \ XCAD \ Xada,M。Robledo,Primpol:DNA复制酶之间的突破性和潜在的癌症治疗靶标的癌症治疗,生物分子,12(20222)。 [7] A.O. Giacomelli,X。Yang,R.E。 Lintner,J.M。McFarland,M。Duby,J.Kim,T.P。 D.Y. Howard武田Ly,E。Kim,H.S。甘农(B. J.G. Aguirre Doench,F。Piccioni,C.W.M。 Roberts,M。Meyerson,G。Getz,C.M。约翰内森,D.E。根,W.C。 Hahn,突变过程塑造了人类癌症中TP53突变的景观,NAT Genet,50(2018)1381-1387。 [8] G.A. Fontana,H.L。Gahlon,线粒体DNA缺失形成的复制和修复机制,核酸res,48(2020)11244-11258。 [9] C.Y. Dai,C.C。 Ng,G.C.C。 Hung,I。Kirmes,L.A](/simg/f\f6a2bb5f504472f4ca10c1ecebfe7702878cdb55.webp)
b'[2] C. Yan,X。Duanmu,L。Zeng,B。Liu,Z。Song,线粒体DNA:分布,突变和消除,细胞,8(2019)。 [3] F. Liu,D.E。 Sanin,X。Wang,肺癌中的线粒体DNA,实验医学和生物学进展,1038(2017)9-22。 [4] J. Zhang,J。 Jia,M。Junaid,Y.B。 MA,F。Ahmad,Y.F。 Jia,W.G。Li,D.S。Pei,人类DNA2(HDNA2)作为癌症和其他疾病的潜在靶标的作用:系统评价,DNA修复(AMST),59(2017)9-19。 [6] A. D \ XC3 \ Xadaz-Talavera,C。Montero-Conde,L.J。Leandro-Garc \ XC3 \ XCAD \ Xada,M。Robledo,Primpol:DNA复制酶之间的突破性和潜在的癌症治疗靶标的癌症治疗,生物分子,12(20222)。 [7] A.O. Giacomelli,X。Yang,R.E。 Lintner,J.M。McFarland,M。Duby,J.Kim,T.P。 D.Y. Howard武田Ly,E。Kim,H.S。甘农(B. J.G. Aguirre Doench,F。Piccioni,C.W.M。 Roberts,M。Meyerson,G。Getz,C.M。约翰内森,D.E。根,W.C。 Hahn,突变过程塑造了人类癌症中TP53突变的景观,NAT Genet,50(2018)1381-1387。 [8] G.A. Fontana,H.L。Gahlon,线粒体DNA缺失形成的复制和修复机制,核酸res,48(2020)11244-11258。 [9] C.Y. Dai,C.C。 Ng,G.C.C。 Hung,I。Kirmes,L.A
b'[2] C. Yan,X。Duanmu,L。Zeng,B。Liu,Z。歌曲,线粒体DNA:分布,突变和消除,细胞,8(2019)。[3] F. Liu,D.E。Sanin,X。Wang,肺癌中的线粒体DNA,实验医学与生物学进展,1038(2017)9-22。[4] J. Zhang,J。[5] P.P.Jia,M。Junaid,Y.B。 MA,F。Ahmad,Y.F。 jia,W.G。 li,D.S。 pei,人类DNA2(HDNA2)作为癌症和其他疾病的潜在靶点的作用:系统评价,DNA修复(AMST),59(2017)9-19。 [6] A. D \ XC3 \ Xadaz-Talavera,C。Montero-Conde,L.J。 Leandro-Garc \ XC3 \ Xada,M。Robledo,Primpol:DNA复制酶的突破和潜在的癌症治疗新靶标,生物分子,12(2022)。 [7] A.O. Giacomelli,X。Yang,R.E。 lintner,J.M. McFarland,M。Duby,J。Kim,T.P。 D.Y. Howard Takeda,S.H。 ly,E。Kim,H.S。 Gannon,B。Hurhula,T。Sharpe,A。Goodale,B。Fritchman,S。Seelman,F。Vazquez,A。Tsherniak,A.J。 Aguirre,J.G。 Doench,F。Piccioni,C.W.M。 Roberts,M。Meyerson,G。Getz,C.M。 Johannessen,D.E。 根,W.C。 Hahn,突变过程塑造了人类癌症中TP53突变的景观,NAT Genet,50(2018)1381-1387。 [8] G.A. Fontana,H.L。 [9] C.Y. dai,C.C。 ng,G.C.C。 Hung,I。Kirmes,L.A。Hughes,Y。Jia,M。Junaid,Y.B。MA,F。Ahmad,Y.F。 jia,W.G。 li,D.S。 pei,人类DNA2(HDNA2)作为癌症和其他疾病的潜在靶点的作用:系统评价,DNA修复(AMST),59(2017)9-19。 [6] A. D \ XC3 \ Xadaz-Talavera,C。Montero-Conde,L.J。 Leandro-Garc \ XC3 \ Xada,M。Robledo,Primpol:DNA复制酶的突破和潜在的癌症治疗新靶标,生物分子,12(2022)。 [7] A.O. Giacomelli,X。Yang,R.E。 lintner,J.M. McFarland,M。Duby,J。Kim,T.P。 D.Y. Howard Takeda,S.H。 ly,E。Kim,H.S。 Gannon,B。Hurhula,T。Sharpe,A。Goodale,B。Fritchman,S。Seelman,F。Vazquez,A。Tsherniak,A.J。 Aguirre,J.G。 Doench,F。Piccioni,C.W.M。 Roberts,M。Meyerson,G。Getz,C.M。 Johannessen,D.E。 根,W.C。 Hahn,突变过程塑造了人类癌症中TP53突变的景观,NAT Genet,50(2018)1381-1387。 [8] G.A. Fontana,H.L。 [9] C.Y. dai,C.C。 ng,G.C.C。 Hung,I。Kirmes,L.A。Hughes,Y。MA,F。Ahmad,Y.F。jia,W.G。li,D.S。pei,人类DNA2(HDNA2)作为癌症和其他疾病的潜在靶点的作用:系统评价,DNA修复(AMST),59(2017)9-19。[6] A. D \ XC3 \ Xadaz-Talavera,C。Montero-Conde,L.J。Leandro-Garc \ XC3 \ Xada,M。Robledo,Primpol:DNA复制酶的突破和潜在的癌症治疗新靶标,生物分子,12(2022)。[7] A.O.Giacomelli,X。Yang,R.E。 lintner,J.M. McFarland,M。Duby,J。Kim,T.P。 D.Y. Howard Takeda,S.H。 ly,E。Kim,H.S。 Gannon,B。Hurhula,T。Sharpe,A。Goodale,B。Fritchman,S。Seelman,F。Vazquez,A。Tsherniak,A.J。 Aguirre,J.G。 Doench,F。Piccioni,C.W.M。 Roberts,M。Meyerson,G。Getz,C.M。 Johannessen,D.E。 根,W.C。 Hahn,突变过程塑造了人类癌症中TP53突变的景观,NAT Genet,50(2018)1381-1387。 [8] G.A. Fontana,H.L。 [9] C.Y. dai,C.C。 ng,G.C.C。 Hung,I。Kirmes,L.A。Hughes,Y。Giacomelli,X。Yang,R.E。lintner,J.M.McFarland,M。Duby,J。Kim,T.P。 D.Y. Howard Takeda,S.H。 ly,E。Kim,H.S。 Gannon,B。Hurhula,T。Sharpe,A。Goodale,B。Fritchman,S。Seelman,F。Vazquez,A。Tsherniak,A.J。 Aguirre,J.G。 Doench,F。Piccioni,C.W.M。 Roberts,M。Meyerson,G。Getz,C.M。 Johannessen,D.E。 根,W.C。 Hahn,突变过程塑造了人类癌症中TP53突变的景观,NAT Genet,50(2018)1381-1387。 [8] G.A. Fontana,H.L。 [9] C.Y. dai,C.C。 ng,G.C.C。 Hung,I。Kirmes,L.A。Hughes,Y。McFarland,M。Duby,J。Kim,T.P。D.Y. Howard Takeda,S.H。 ly,E。Kim,H.S。 Gannon,B。Hurhula,T。Sharpe,A。Goodale,B。Fritchman,S。Seelman,F。Vazquez,A。Tsherniak,A.J。 Aguirre,J.G。 Doench,F。Piccioni,C.W.M。 Roberts,M。Meyerson,G。Getz,C.M。 Johannessen,D.E。 根,W.C。 Hahn,突变过程塑造了人类癌症中TP53突变的景观,NAT Genet,50(2018)1381-1387。 [8] G.A. Fontana,H.L。 [9] C.Y. dai,C.C。 ng,G.C.C。 Hung,I。Kirmes,L.A。Hughes,Y。D.Y. HowardTakeda,S.H。 ly,E。Kim,H.S。 Gannon,B。Hurhula,T。Sharpe,A。Goodale,B。Fritchman,S。Seelman,F。Vazquez,A。Tsherniak,A.J。 Aguirre,J.G。 Doench,F。Piccioni,C.W.M。 Roberts,M。Meyerson,G。Getz,C.M。 Johannessen,D.E。 根,W.C。 Hahn,突变过程塑造了人类癌症中TP53突变的景观,NAT Genet,50(2018)1381-1387。 [8] G.A. Fontana,H.L。 [9] C.Y. dai,C.C。 ng,G.C.C。 Hung,I。Kirmes,L.A。Hughes,Y。Takeda,S.H。ly,E。Kim,H.S。Gannon,B。Hurhula,T。Sharpe,A。Goodale,B。Fritchman,S。Seelman,F。Vazquez,A。Tsherniak,A.J。 Aguirre,J.G。 Doench,F。Piccioni,C.W.M。 Roberts,M。Meyerson,G。Getz,C.M。 Johannessen,D.E。 根,W.C。 Hahn,突变过程塑造了人类癌症中TP53突变的景观,NAT Genet,50(2018)1381-1387。 [8] G.A. Fontana,H.L。 [9] C.Y. dai,C.C。 ng,G.C.C。 Hung,I。Kirmes,L.A。Hughes,Y。Gannon,B。Hurhula,T。Sharpe,A。Goodale,B。Fritchman,S。Seelman,F。Vazquez,A。Tsherniak,A.J。Aguirre,J.G。 Doench,F。Piccioni,C.W.M。 Roberts,M。Meyerson,G。Getz,C.M。 Johannessen,D.E。 根,W.C。 Hahn,突变过程塑造了人类癌症中TP53突变的景观,NAT Genet,50(2018)1381-1387。 [8] G.A. Fontana,H.L。 [9] C.Y. dai,C.C。 ng,G.C.C。 Hung,I。Kirmes,L.A。Hughes,Y。Aguirre,J.G。Doench,F。Piccioni,C.W.M。Roberts,M。Meyerson,G。Getz,C.M。 Johannessen,D.E。 根,W.C。 Hahn,突变过程塑造了人类癌症中TP53突变的景观,NAT Genet,50(2018)1381-1387。 [8] G.A. Fontana,H.L。 [9] C.Y. dai,C.C。 ng,G.C.C。 Hung,I。Kirmes,L.A。Hughes,Y。Roberts,M。Meyerson,G。Getz,C.M。Johannessen,D.E。 根,W.C。 Hahn,突变过程塑造了人类癌症中TP53突变的景观,NAT Genet,50(2018)1381-1387。 [8] G.A. Fontana,H.L。 [9] C.Y. dai,C.C。 ng,G.C.C。 Hung,I。Kirmes,L.A。Hughes,Y。Johannessen,D.E。根,W.C。 Hahn,突变过程塑造了人类癌症中TP53突变的景观,NAT Genet,50(2018)1381-1387。[8] G.A.Fontana,H.L。 [9] C.Y. dai,C.C。 ng,G.C.C。 Hung,I。Kirmes,L.A。Hughes,Y。Fontana,H.L。[9] C.Y.dai,C.C。ng,G.C.C。Hung,I。Kirmes,L.A。Hughes,Y。gahlon,线粒体DNA缺失形成的复制和修复机制,核酸res,48(2020)11244-11258。du,C.A。Brosnan,A。Ahier,A。Hahn,C.M。 Haynes,O。Rackham,A。Filipovska,S。Zuryn,ATFS-1,通过促进转录修复,自然细胞生物学,25(2023)1111-1120来抵消线粒体DNA损伤。 [10] L. Ou,H。Liu,C。Peng,Y. [11] H. Liu,J。Weng,C.L.H。 Huang,A.P。 杰克逊,癌症的电压门控钠通道,生物标志物研究,12(2024)70。 [12] H. Liu,A。Dong,A.M。 Rasteh,P。Wang,J。Weng,乳腺癌中新型T细胞CD8 +标记的鉴定,Scientific Reports,14(2024)19142。 [13] H. Liu,T。Tang,基于MAPK信号途径的胶质瘤亚型,机器学习风险模型和关键集线器蛋白识别,科学报告,13(2023)19055。。 [14] H. Liu,T。Tang,《泛滥成灾与基因集的泛癌遗传分析》,癌症遗传学,278-279(2023)91-103。 [15] H. Liu,T。Tang,《胶质瘤IGFBP的生物信息学研究》,涉及其诊断,预后和治疗预测值,AM J Transl Res,15(2023)2140-2155。 [16] H. Liu,T。Tang,《泛滥成灾基因套件的泛 - 癌遗传分析》,Biorxiv,(2023),2023.2002。 2025.529997。 [17] H. Liu,库糖凋亡在肾脏肾透明细胞癌中的表达和潜在免疫受累,癌症遗传学,274-275(2023)21-25。Brosnan,A。Ahier,A。Hahn,C.M。Haynes,O。Rackham,A。Filipovska,S。Zuryn,ATFS-1,通过促进转录修复,自然细胞生物学,25(2023)1111-1120来抵消线粒体DNA损伤。[10] L. Ou,H。Liu,C。Peng,Y.[11] H. Liu,J。Weng,C.L.H。Huang,A.P。 杰克逊,癌症的电压门控钠通道,生物标志物研究,12(2024)70。 [12] H. Liu,A。Dong,A.M。 Rasteh,P。Wang,J。Weng,乳腺癌中新型T细胞CD8 +标记的鉴定,Scientific Reports,14(2024)19142。 [13] H. Liu,T。Tang,基于MAPK信号途径的胶质瘤亚型,机器学习风险模型和关键集线器蛋白识别,科学报告,13(2023)19055。。 [14] H. Liu,T。Tang,《泛滥成灾与基因集的泛癌遗传分析》,癌症遗传学,278-279(2023)91-103。 [15] H. Liu,T。Tang,《胶质瘤IGFBP的生物信息学研究》,涉及其诊断,预后和治疗预测值,AM J Transl Res,15(2023)2140-2155。 [16] H. Liu,T。Tang,《泛滥成灾基因套件的泛 - 癌遗传分析》,Biorxiv,(2023),2023.2002。 2025.529997。 [17] H. Liu,库糖凋亡在肾脏肾透明细胞癌中的表达和潜在免疫受累,癌症遗传学,274-275(2023)21-25。Huang,A.P。杰克逊,癌症的电压门控钠通道,生物标志物研究,12(2024)70。[12] H. Liu,A。Dong,A.M。 Rasteh,P。Wang,J。Weng,乳腺癌中新型T细胞CD8 +标记的鉴定,Scientific Reports,14(2024)19142。[13] H. Liu,T。Tang,基于MAPK信号途径的胶质瘤亚型,机器学习风险模型和关键集线器蛋白识别,科学报告,13(2023)19055。[14] H. Liu,T。Tang,《泛滥成灾与基因集的泛癌遗传分析》,癌症遗传学,278-279(2023)91-103。[15] H. Liu,T。Tang,《胶质瘤IGFBP的生物信息学研究》,涉及其诊断,预后和治疗预测值,AM J Transl Res,15(2023)2140-2155。[16] H. Liu,T。Tang,《泛滥成灾基因套件的泛 - 癌遗传分析》,Biorxiv,(2023),2023.2002。2025.529997。[17] H. Liu,库糖凋亡在肾脏肾透明细胞癌中的表达和潜在免疫受累,癌症遗传学,274-275(2023)21-25。[18] L. Hengrui,《中药用于癌症治疗中使用的有毒药物的例子》,J Tradit Chin Med,43(2023)209-210。[19] H. Liu,J。Weng,《 Rad51的Pan-Cancer生物信息学分析》,涉及诊断,预后和治疗预测的值,肿瘤学的前沿,12(2022)。[20] H. Liu,J。Weng,胶质瘤中细胞周期蛋白依赖性激酶2(CDK2)的全面生物信息学分析,Gene,(2022)146325。[21] H. Liu,T。Tang,Pan-Cancer的库糖胞化和铜代谢相关的基因集,肿瘤学的边界,12(2022)952290。[22] H. Liu,Y。Li,Cornichon家族AMPA受体辅助蛋白4(CNIH4)在头部和颈部鳞状细胞癌中的潜在作用,癌症生物标志物:疾病标志物A部分(2022)。[23] H. Liu,J.P。Dilger,J。Lin,pan-Cancer-Biodorminicals-Informinical-Informicals Trpm7的文献综述,Pharmacol Ther(2022)108302。[24] H. Liu,cuproptosis Gene Set的Pan-Canter概况,《美国癌症研究杂志》,第12期(2022)4074-4081。[25] Y. Liu,H。Liu,氨基酰基TRNA合成酶复合物的临床能力相互作用多功能蛋白1(AIMP1),用于头颈鳞状细胞癌,癌症生物标志物:疾病标志物A节A节(20222)。[26] Y. Li,H。Liu,Y。Han,在头部和颈部鳞状细胞癌中,Cornichon家族AMPA受体辅助蛋白4(CNIH4)的潜在作用,研究方形(2021)。 '
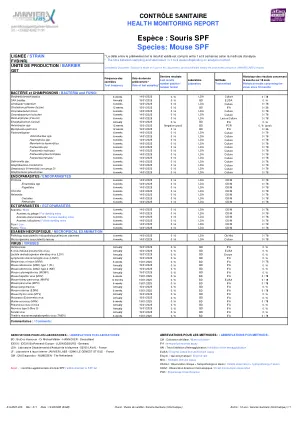
物种:SPF鼠标
Bactérieet Champignons /细菌和Fung Bonchiseptica 6周14/01/2025 0 /6 LDA文化Rodentum 6周14/01/2025 0/6 LDA文化0/78 Clostrid Piliforme(Tyzzer)12周15/01/2025 0/6 BDFAA 0/36 BOVIS BOVIS BOVIS 6周14/01/2025 0/2025 0/6 6 LDA培养0 /78 Cownebacterium Kutscheri 6周14/01/2025 0 /6 LDA培养0/78皮肤植物(IIF病变)年度15/01/2025 0/6 BD灯0/6 Helicabacter spp 12周15/01/2025负(池)BD 0/6(POL)Mycoplasma肺部12周15/01/2025 0/6 BD IFAA IFAA 0/ 36巴氏菌科6周14/01/2025 0 /6 LDA培养0/78 actinobcillus spp。6周14/01/2025 0 /6 LDA文化0/78 Heamophilus spp。6周14/01/2025 0 /6 LDA培养0/78 Mannheimia Haemytica Haemytica 6周14/01/2025 0 /6 LDA培养0/78 Pasteurella spp。6周14/01/2025 0/6 LDA培养0/78糊状multocida 6周14/01/2025 0/6 LDA培养0/78 pneumotropica pneumotropica 6 Weeks 14/01/2025 0/6 LDA LDA培养0 /78 treehalosi的巴斯德奶酪6周14/01/2025 0 /6 LDA培养0/78 Salmonnella sp。6 weeks 14/01/2025 0 / 6 LDA Culture 0 / 78 Streptobacillus moniliformis 6 weeks 14/01/2025 0 / 6 LDA Culture 0 / 78 Streptococci ß-hemolytic (not group D) 6 weeks 14/01/2025 0 / 6 LDA Culture 0 / 78 Streptococcus pneumoniae 6 weeks 14/01/2025 0 / 6 LDA Culture 0 / 78 ENDOPARASITES / ENDOPARASITES Protozoa 6 weeks 14/01/2025 0 / 6 LDA OD/M 0 / 78 Entamoeba spp 6 weeks 14/01/2025 0 / 6 LDA OD/M 0 / 78 Flagellates 6 weeks 14/01/2025 0 / 6 LDA OD/M 0 / 78 Coccidia 6 weeks 14/01/2025 0 / 6 LDA OD/M 0 / 78 Helminths 6 weeks 14/01/2025 0 / 6 LDA OD/M 0 / 78 Cestodes 6 weeks 14/01/2025 0 / 6 LDA OD/M 0 / 78 Nematodes 6 weeks 14/01/2025 0 / 6 LDA OD/M 0 / 78 ECTOPARASITES / ECTOPARASITES Acariens / Mites 6 weeks 14/01/2025 0 / 6 LDA OD/M 0 / 78 Acariens du pelage / Fur-dwelling mites 6 weeks 14/01/2025 0 / 6 LDA OD/M 0 / 78 Acariens d'environnement / Surface-dwelling mites 6 weeks 14/01/2025 0 / 6 LDA OD/M 0 / 78 Acariens folliculaires/ Follicle-dwelling mites 6星期14/01/2025 0/6 LDA OD/M 0/78 POUX/LICE 6周14/01/2025 0/6 LDA OD/M 0/M 0/M 0/M 0/78 PUCES/FLEAS 6周14/01/2025 0/6 LDA OD/M 0/M 0/M 0/78 ISCROPSIQIE/NECROPSIQUIQIE/NECROPSIQUE/NECOPIQUES QUSTOPIQUE/NECOPIQUES POOCTOPOPIQUE/NECOPIQUES速度与历史悠久的病理学logation 5 / 6 LDA OB / HIST 0 /78与病变相关的微生物6周14/01/2025 0 /6 LDA培养0/78每年15/01/2025 0 /6 BD IFA 0 /6 K病毒(小鼠肺炎病毒)每年15/6 BD hantaviruses hantaviruses每年15/0 / 6 hhydrase 15/01/201/201/2025 0 /6 BD D.病毒(LDV)每年15/01/2025 0/6 BD酶。0 /6每年15/01/2025 0 /6 BD IFA 0/6分钟病毒(MVM)6周15/01/2025 0 /6 BD IFA IFA 0 /78小鼠腺病毒(MAD)IFA(MAD)IFA 0 /6 BDA 0 /6 BD, (MAD)类型2(K87)每年15/01/2025 0 /6 ifa 0/6鼠标巨细胞性病毒(MCMV)每年15/01/2025 0 /6 BD IFA 0 /6鼠标肝炎病毒病毒(MHV)6周6周15/01/2025 0 /6 BD IFA 0 /78 IFA 0 /78 MOUSS NEDNEY(MOK) 15/01/2025 0 /6 BD ELISA 0/18鼠标细小病毒(MPV)6周15/01/2025 0 /6 BD IFA 0/78鼠标多组病每年15/01/2025 0 /6 BD IFA 0/6 ifa 0/6 6 Weeke Rotavirus(Edim)6周15/01/201/201/2025 0 /6 BD, Annually 15/01/2025 0 / 6 BD IFA 0 / 6 Mousepox (Ectromelia) virus Annually 15/01/2025 0 / 6 BD IFA 0 / 6 Murine norovirus (MNV) 6 weeks 15/01/2025 0 / 6 BD IFA 0 / 78 Pneumonia virus of mice Annually 15/01/2025 0 / 6 BD IFA 0 / 6 Reovirus 3型(REO 3)每年15/01/2025 0/6 BD IFA 0/6每年15/01/2025 0/6 BD IFA 0/6 Theiler的鼠脑静脉炎病毒病毒(TMEV)6周6周15/01/2025 0/6 BD 0/6 BD IFA 0/78 ifa 0/78