XiaoMi-AI文件搜索系统
World File Search SystemMHV
免疫疗法和靶向辐射收缩肝肿瘤,使手术
这种方法允许外科医生快速准确地定位MHV,这是肝脏的主要血管之一。在常规方法中,MHV造成分裂损伤或医源性损害的风险更高,这可能导致严重的并发症,特别是在功能性肝脏储备有限的肝硬化患者中。通过关注这个里程碑,Arantius-Fir-Fir的技术最大程度地减少了伤害MHV并确保其保存的风险,这在肝硬化患者的左肝切除术中至关重要。此精度不仅提高了程序的安全性,还可以有助于更好的患者预后。

机械心脏瓣膜孕妇的抗凝治疗:意大利抗肉眼的诊断和监测中心联合会
摘要机械心脏瓣膜(MHV)孕妇抗凝治疗的管理很难,即使对于领域中经历的临床医生也经常具有挑战性。与没有MHV的女性相比,这些妊娠的负担负担母亲和胎儿的并发症率更高。由维生素K拮抗剂提供的最佳抗凝作用的母体需求与它们对胚胎和胎儿的破坏作用相抵消。另一方面,考虑到这些患者的血栓形成并发症的高风险。因此,有关MHV孕妇管理的许多临床问题仍未得到答复,例如选择最佳的抗凝剂剂,要达到和维持的最佳抗凝水平以及对母亲和胎儿的长期影响的评估。基于对当前文献的全面综述,即诊断和监视抗血栓形成疗法(FCSA)的意大利联合会提出了基于经验的建议和专家意见。尤其是,该共识文件旨在为与患有MHV的孕妇打交道的临床医生提供实用指导,以优化孕产妇和胎儿的结局,同时保证适当的抗癌。最后,FCSA强调了建立在怀孕期间孕妇管理孕妇的多学科团队的必要性,

报告疾病和健康危害
2023年11月11日,19:30,萨达奥的边境控制检查站收到了道路事故的报告,导致一场死亡和五人受伤。所有参与的人都是马来西亚的公民。检查站当局与Bukit Kayu Hitam检查站的马来西亚卫生官员协调,为患者和死者的转移做准备。通过用于两国官员之间的沟通的WhatsApp申请来促进这种协调。操作和文档验证顺利进行。2023年11月12日,泰国世界宣明会基金会在泰国世界卫生组织的支持下,在曼谷的Ramakarndeen Hotel组织了一次移民卫生志愿者(MHV)培训研讨会。22个MHV来自曼谷和Pathum Thani。目的是为志愿者提供有关疾病预防和控制的准确信息。重点是对以下关键疾病的监视和管理:禽流感和登革热。的目的是使MHV能够在各自的责任领域有效地传播知识,尤其是在预防和控制疾病暴发方面。


2024-2025 学年学校教育计划
为所有学生提供各种课程和学习,包括:将基于土地的学习日纳入课程成果,与梅蒂斯长老艺术家建立持续关系,担任 MHV 驻校艺术家,使用第一民族梅蒂斯和因纽特套件(传统土著游戏单元、巨型地图)和七大神圣教义作为品格教育计划的一部分。全年,EIPS 顾问将为员工举办专业学习,以增加基础知识,以便教师可以为每个班级创建土地认知,并创建用于基于土地的学习的自然手推车。

研究NSP15氨基末端结构域在NSP15功能,病毒复制和发病机理中的作用
图19。筛选病毒感染期间的内向功能丧失...........................................................................................................................................................................................................................................................................................................................................................................................................................................................................................................................................利用原位HA标签的NSP15病毒...........................................................................................................................................................................................................................................................................................................................................................................................................................Interactions Between Nsp15 and RTC Co-factor Nsp8 were Not Disrupted by the E3K Mutation ........................................................................................................... 64 Figure 22.MHV-A59 NSP15六聚体结构的模型突出了L2和E3的氢键网络。 .................................................................... 65 Figure 23. NTD Mutations Do Not Attenuate Viral Replication in Non-interferon Responsive BMDMs ...................................................................................................... 66 Figure 24. MHV NTD突变病毒在BMDMS感染期间积累了类似WT的NSP15蛋白水平。 MHV NTD Mutants are Attenuated in BMDMs and Induce a Robust Interferon Response ....................................................................................................................... 68 Figure 26. Disruption of the Nsp15 NTD Leads to an Early, Robust Activation of OAS/RNaseL Signaling .................................................................................................. 69 Figure 27. SARS-COV-2 NSP15 NTD的突变减弱了IAT2中的病毒复制。 ............................................................................................................................. 70 Figure 28.MHV-A59 NSP15六聚体结构的模型突出了L2和E3的氢键网络。.................................................................... 65 Figure 23.NTD Mutations Do Not Attenuate Viral Replication in Non-interferon Responsive BMDMs ...................................................................................................... 66 Figure 24.MHV NTD突变病毒在BMDMS感染期间积累了类似WT的NSP15蛋白水平。MHV NTD Mutants are Attenuated in BMDMs and Induce a Robust Interferon Response ....................................................................................................................... 68 Figure 26.Disruption of the Nsp15 NTD Leads to an Early, Robust Activation of OAS/RNaseL Signaling .................................................................................................. 69 Figure 27.SARS-COV-2 NSP15 NTD的突变减弱了IAT2中的病毒复制。 ............................................................................................................................. 70 Figure 28.SARS-COV-2 NSP15 NTD的突变减弱了IAT2中的病毒复制。............................................................................................................................. 70 Figure 28.SARS-COV-2 NSP15 NTD突变体在IAT2中诱导早期,稳健的ISG表达。............................................................................................................................. 71 Figure 29.丧失内向活性的丧失会引起感染BMDMS的转录组轮廓的急剧变化。............................................................................................ 72 Figure 30.NSP15突变病毒在BMDM感染过程中诱导了几种IFN和DSRNA传感器基因的表达。......................................................................... 73 Figure 31.在NSP15突变病毒感染期间,参与坏死途径的基因被上调。................................................................................................... 73 Figure 32.NSP15突变病毒诱导ZBP1依赖性坏死性................................................................................................................................................................................... 74图33。Nsp15 Mutant Viruses Induce ZBP1-independent Apoptosis and Necroptosis ....................................................................................................................................... 76 Figure 34.C57BL/6 Mice Infected with Nsp15 NTD Mutant Viruses Do Not Lose Weight ....................................................................................................................................... 77

360 Diagnostics™:2020 US目录|查尔斯河 - 网
鼠标细胞视角病毒(MVM/MPV)••••••••••••••••••••••••••••••••••••••••••••••••••••••••••••••••••••••••••••••••••••••••••。 Adenovirus type 1 & 2 (MAV-1 & MAV-2) • • • Reovirus type 1, 2, 3, 4 • • • Pneumonia virus of mice • • Sendai virus • • Ectromelia (mousepox) • • • Lymphocytic choriomeningitis virus • • • Astrovirus 1 & 2 New World hantavirus † Bacteria Helicobacter • • • • • • •柠檬杆菌•••••••••••••••••••••••••••••••••••••••••••••••••••••••••••••••••••••••••••••••••••••••弯曲杆菌•••••••••••••••••••••••••••••••••••••••••••••••••••••••••••••••••••••••••••••••••••••••••••••••••••••••••••••••••••• Oxytoca•••••β溶血性链球菌A•••••β溶血性链球菌B组B•β溶血性链球菌C组C组C•••••••••••••••••••••••••••••••••••••••••••••••••••••••••••••••••••••••••••••••••••••••••••••••••••••••••••••••。

RNA病毒克隆平台 有历史的小鼠
《自然》杂志的一项研究报告了酵母酿酒酵母作为组装和维护各种 RNA 病毒基因组(包括 SARS-CoV-2)的平台的适用性,该平台可实现对 SARS-CoV-2 的基因操作和功能表征。在疫情爆发期间,病毒分离株可用于开发诊断、体内模型、抗病毒疗法和疫苗。如果病毒分离株的可用性有限,可以从化学合成的 DNA 中克隆病毒基因组,但使用大肠杆菌的既定方法通常不足以容纳冠状病毒(冠状病毒科)等 RNA 病毒的大型基因组。Thao 等人将转化相关重组 (TAR) 克隆应用于含有 GFP 基因的小鼠肝炎病毒 (MHV),该病毒具有成熟的反向遗传学平台。将覆盖 MHV-GFP 基因组和 TAR 载体的重叠 DNA 片段转化到酵母中,DNA 片段通过同源重组组装,产生包含全长病毒 cDNA 的酵母人工染色体 (YAC)。值得注意的是,90% 以上的筛选克隆显示 YAC 组装正确,表明组装效率高。通过分离和线性化 YAC 进行体外转录以生成病毒 RNA,成功从两个单个克隆中回收了传染性病毒,然后将其与编码 MHV 核衣壳蛋白的 mRNA 一起转染到 BHK-MHV-N 仓鼠细胞系中,以产生和扩增病毒。回收的病毒表现出与亲本 MHV-GFP 相同的复制动力学。该团队着手确定合成基因组学平台是否可以应用于 MERS-CoV,使用低拷贝细菌人工染色体 (BAC) 从八个重叠的 PCR 扩增 DNA 片段克隆病毒。该方法还应用于突变的 MERS-CoV 克隆,该克隆中插入了 GFP 基因。YAC 克隆组装和从克隆 DNA 中拯救病毒均取得成功,确定了该平台可适用于更广泛的病毒,包括转基因病毒基因组。进一步的实验确定病毒基因组可以稳定维持,并且该平台适用于其他难以克隆的病毒,例如寨卡病毒(黄病毒科)和人类呼吸道合胞病毒(副粘病毒科),包括直接从临床样本中克隆,而无需事先了解病毒基因型。令人惊讶的是,在收到基于 2020 年 1 月发布的基因组序列的 SARS-CoV-2 合成 DNA 片段后 1 周内,就实现了重组 SARS-CoV-2 和 SARS-CoV-2-GFP 的克隆和拯救。总之,这项研究展示了合成基因组学平台在疫情期间从不同起始材料(包括病毒分离物、克隆 DNA、合成 DNA 或临床样本)快速生成和功能表征进化 RNA 病毒的实用性。

病毒编码的蛋白酶和Nidovirales中的蛋白水解加工
基于基因组结构和复制策略的相似性,RNA病毒如今可分为“超类群”,通常涵盖动物病毒和植物病毒(Goldbach & Wellink,1988;Strauss & Strauss,1988)。这一概念也越来越多地体现在病毒分类学中;尤其是引入了分类单元“目”,将很可能拥有共同祖先的病毒科合并在一起(Mayo & Pringle,1998)。对于正链、有包膜的冠状病毒和动脉炎病毒(最近被统一归入巢病毒目,Cavanagh,1997),基于相似的多顺反子基因组结构、共同的转录和(后)翻译策略以及一系列同源复制酶结构域的保守性(den Boon et al.,1991),它们之间建立了密切的系统发育关系。因此,有可能勾勒出nidovirus生命周期的共同轮廓(图1)(详见Lai & Cavanagh,1997;de Vries et al.,1997;Snijder & Meulenberg,1998)。然而,在某些方面,这两个病毒家族彼此之间存在显著差异。例如,最大的冠状病毒基因组,鼠肝炎病毒(MHV),其基因组为31±5kb,约为最小动脉炎病毒基因组,即马动脉炎病毒(EAV)12±7kb RNA的两倍半。此外,这两个病毒家族的结构蛋白没有明显的相关性,导致病毒体的大小和结构存在重要差异(den Boon et al.,1991;Snijder & Spaan,1995;de Vries et al.,1997)。大多数主要的动物正链RNA病毒群体要么产生单个多聚蛋白,要么产生单独的非结构和结构前体多肽,这些多肽随后被病毒编码或宿主编码的蛋白酶裂解,产生功能性亚基(Dougherty & Semler, 1993)。相比之下,在基因组3′-近端区域编码的nido病毒结构蛋白,
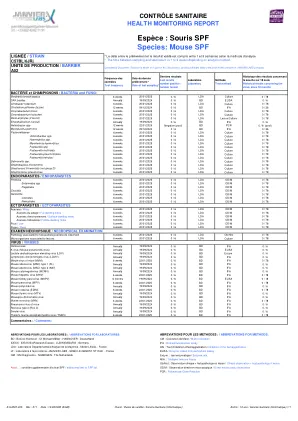
物种:SPF鼠标
Bactérieet Champignons /细菌和Fung Bonchiseptica 6周20/01/2025 0 /6 LDA文化Rodentium 6周20/01/2025 0/6 LDA文化0/78梭状芽胞杆菌(Tyzzer)12周09/12/2024 0/6 BD IFA 0/36 BOVIS BOVIS BOVIS 6周20/01/2025 0/2025 0/2025 0/6 LDA培养0 /78 Cownebacterium Kutscheri 6周20/01/2025 0 /6 LDA培养0/78皮肤植物(如果病变)年度19/09/2024 0/6 BD灯0/6 Helicabacter SPP 12周22/01/2025负(POLOL)BD 0/6(POL)Mycoplasma plemonis 12 Weeks 09/12/2024 0/6 BD IFAA 0/6 BD IFAA 0/ 36巴氏菌科6周20/01/2025 0 /6 LDA培养0/78 actinobcillus spp。6周20/01/2025 0 /6 LDA文化0/78 Heamophilus spp。6周20/01/2025 0 /6 LDA培养0/78 Mannheimia Haemytica Haemytica 6周20/01/2025 0 /6 LDA培养0/78 Pasteurella spp。6周20/01/2025 0/6 LDA培养0/78糊剂Multocida 6周20/01/2025 0/6 LDA培养0/78 pneumotropica pereumotropica 6 Weeks 20/01/2025 0/6 LDA LDA培养0 /78 treehalosi的巴斯德奶酪6周20/01/2025 0 /6 LDA培养0/78 Salmonnella spp。6 weeks 20/01/2025 0 / 6 LDA Culture 0 / 78 Streptobacillus moniliformis 6 weeks 20/01/2025 0 / 6 LDA Culture 0 / 78 Streptococci ß-hemolytic (not group D) 6 weeks 20/01/2025 0 / 6 LDA Culture 0 / 78 Streptococcus pneumoniae 6 weeks 20/01/2025 0 / 6 LDA Culture 0 / 78 ENDOPARASITES / ENDOPARASITES Protozoa 6 weeks 20/01/2025 0 / 6 LDA OD/M 0 / 78 Entamoeba spp 6 weeks 20/01/2025 0 / 6 LDA OD/M 0 / 78 Flagellates 6 weeks 20/01/2025 0 / 6 LDA OD/M 0 / 78 Coccidia 6 weeks 20/01/2025 0 / 6 LDA OD/M 0/78蠕虫6周20/01/2025 0/6 LDA OD/M 0/78 CESTODES 6周6周20/01/2025 0/6 LDA OD/M 0/M 0/M 0/M 0/78 NEMATODES 6 WEDWEN 20/01/01/01/2025 0/6 LDA OD/M 0/M 0/M 0/M 0/78 MIMESITES 6周6周6周6周6周20/01/2025 0/6 LDA OD/ M 0/78 Acariens du pelage/ tur-dewelling螨虫6周20/01/2025 0/6 LDA OD/ M 0/ M 0/ M 0/78 Acariens d'Acariens d'evironnement/ Surface-D'Erviration/ Surface-Deblist-Dewelling Mecites 6 Weeking 6 Weech weeks 20/01/2025 0 / 6 LDA OD/M 0 / 78 Poux / Lice 6 weeks 20/01/2025 0 / 6 LDA OD/M 0 / 78 Puces / Fleas 6 weeks 20/01/2025 0 / 6 LDA OD/M 0 / 78 EXAMEN NÉCROPSIQUE / NECROPSICAL EXAMINATION Pathology associated to histopathological lesions observed 6 weeks 20/01/2025 0 / 6 LDA OB / HIST 0 /78与病变相关的微生物6周20/01/2025 0 /6 LDA培养0/78每年19/09/2024 0 /6 BD IFA 0 /6 K病毒(小鼠肺炎病毒)每年19/6 k病毒hantaviruess hantaviruses每年19/09/09/09/20/20/2024 0 /6 BD D.病毒(LDV)每年19/09/2024 0/6 BD酶。0 /6每年19/09/2024每年19/09/0 / 6 ifa IFA 0/6分钟病毒(MVM)6周22/01/2025 0 /6 BD IFA IFA 0 /78小鼠adenovirus(MAD)IFA(MAD)IFA IFA IFA IFA EDA IFA IFA 0 /6 BDA 0 /6 BDA 0 /6 BDA 0 /6 BD ANEN / 6 BDIRIUL (MAD)类型2(K87)每年19/09/2024 0 /6 bd IFA 0/6鼠标cytomegalovirus(MCMV)每年19/09/2024 0 /6 BD IFA 0 /6 IFA 0 /6鼠标肝炎病毒病毒(MHV)6周6周22/01/2025 0 /6 BD IFA 0 /78 IFA 0 /78 MOUSS NEDNEY(MOK) 19/09/2024 0 /6 BD ELISA 0/18鼠标细小病毒(MPV)6周6周22/01/2025 0 /6 BD IFA 0/78鼠标多组病每年19/09/2024 0 /6 BD IFA 0/6 ifa 0 /6 Mouse Rotavirus(Edim)6周22/01/201/201/2025 0 /6 BD Annually 19/09/2024 0 / 6 BD IFA 0 / 6 Mousepox (Ectromelia) virus Annually 19/09/2024 0 / 6 BD IFA 0 / 6 Murine norovirus (MNV) 6 weeks 22/01/2025 0 / 6 BD IFA 0 / 78 Pneumonia virus of mice Annually 19/09/2024 0 / 6 BD IFA 0 / 6 Reovirus 3型(REO 3)每年19/09/2024 0/6 BD IFA 0/6每年19/09/2024 0/6 BD IFA 0/6 Theiler的鼠脑瘤瘤病毒病毒(TMEV)6周6周22/01/2025 0/6 BD IFA 0/6 BD IFA 0/78