机构名称:
¥ 2.0
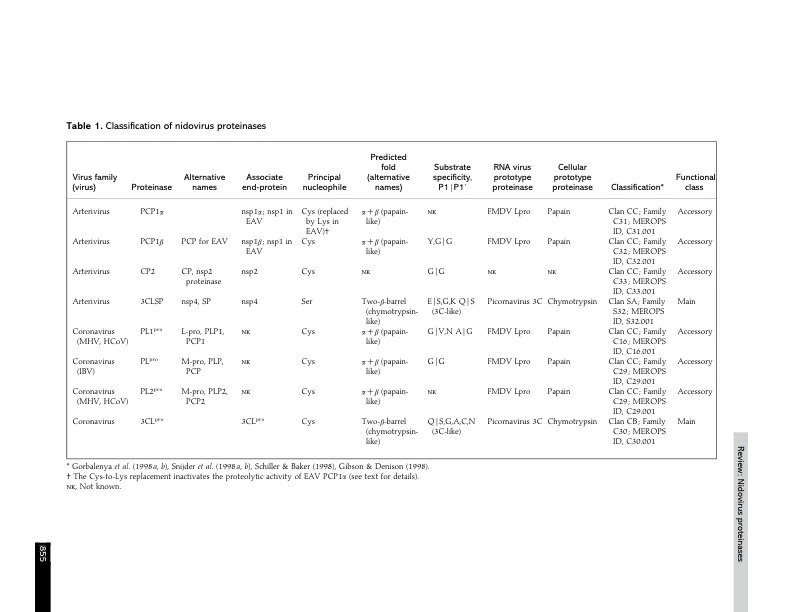
基于基因组结构和复制策略的相似性,RNA病毒如今可分为“超类群”,通常涵盖动物病毒和植物病毒(Goldbach & Wellink,1988;Strauss & Strauss,1988)。这一概念也越来越多地体现在病毒分类学中;尤其是引入了分类单元“目”,将很可能拥有共同祖先的病毒科合并在一起(Mayo & Pringle,1998)。对于正链、有包膜的冠状病毒和动脉炎病毒(最近被统一归入巢病毒目,Cavanagh,1997),基于相似的多顺反子基因组结构、共同的转录和(后)翻译策略以及一系列同源复制酶结构域的保守性(den Boon et al.,1991),它们之间建立了密切的系统发育关系。因此,有可能勾勒出nidovirus生命周期的共同轮廓(图1)(详见Lai & Cavanagh,1997;de Vries et al.,1997;Snijder & Meulenberg,1998)。然而,在某些方面,这两个病毒家族彼此之间存在显著差异。例如,最大的冠状病毒基因组,鼠肝炎病毒(MHV),其基因组为31±5kb,约为最小动脉炎病毒基因组,即马动脉炎病毒(EAV)12±7kb RNA的两倍半。此外,这两个病毒家族的结构蛋白没有明显的相关性,导致病毒体的大小和结构存在重要差异(den Boon et al.,1991;Snijder & Spaan,1995;de Vries et al.,1997)。大多数主要的动物正链RNA病毒群体要么产生单个多聚蛋白,要么产生单独的非结构和结构前体多肽,这些多肽随后被病毒编码或宿主编码的蛋白酶裂解,产生功能性亚基(Dougherty & Semler, 1993)。相比之下,在基因组3′-近端区域编码的nido病毒结构蛋白,
病毒编码的蛋白酶和Nidovirales中的蛋白水解加工
主要关键词




相关文件推荐