XiaoMi-AI文件搜索系统
World File Search SystemNRG1

神经调节蛋白 1 基因 (Nrg1)。一种潜在的治疗肺癌的新型靶向改变
简单总结:肿瘤治疗已经并将继续演变为一种不可知论的方法,即治疗更多地侧重于识别和靶向基因异常,而不是像几十年前那样侧重于癌症的来源器官。随着每种基因异常都被确定为靶点,针对这些基因的药物开发也随之增长,从而提高了生存率和生活质量,人们对寻找新靶点的兴趣也随之增加。肺癌是最好的例子之一,与接受经验性常规化疗的患者相比,可靶向的基因异常导致生存率存在显著差异。神经调节蛋白 1 基因 (NRG1) 的易位是众多具有临床意义的基因融合之一,它有可能成为可靶向的基因,欧洲和美国已经在进行临床试验。本综述旨在描述这种新融合在肺癌治疗中的重要性和最新进展。

晚期非小细胞肺癌 (NSCLC) 中的 NTRK 和 NRG1 基因融合
随着个性化医疗的兴起,非小细胞肺癌也开始出现。自 2004 年发现表皮生长因子受体 (EGFR) 突变以来,从分子水平上定义的可从靶向治疗中获益的患者亚组名单已大幅增加,目前的国际指南建议对所有新诊断的局部晚期或转移性非鳞状非小细胞肺癌患者进行至少 5-8 种生物标志物的分子检测,以选择最佳患者 (5-9)。非小细胞肺癌中 ALK 重排的惊人故事促使人们寻找其他可能用于靶向治疗的致癌重排。神经调节蛋白 - 1 (NRG1) 和 NTRK 融合是最近在非小细胞肺癌中发现的两种重排,是肿瘤不可知生物标志物的两个杰出例子。虽然这两种基因异常相对罕见,但它们代表了 NSCLC 的两个临床相关亚组,可以从靶向治疗中获益。本文我们全面概述了 NRG1 和 NTRK 重排 NSCLC 的生物学和临床病理学特征,以及关于利用这些靶点进行治疗的现有数据。

Bizengri批准的Merus新闻稿
Merus宣布NRG1 +胰腺腺癌和NRG1 + NRG1 +非Small细胞肺癌(NSCLC)的FDA批准(Zenocutuzumab-ZBCO)基于Enrgy研究的安全性和有效性数据
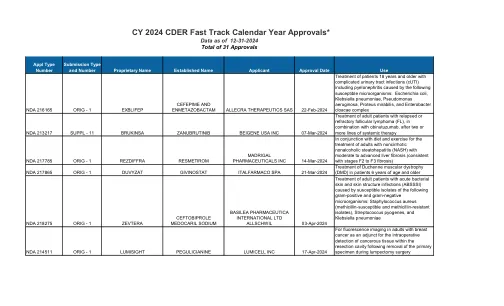
CY 2024 CDER快速日历年批准*
对具有神经调节蛋白1(NRG1)基因融合的晚期,无法切除或转移性非小细胞肺癌的成年人进行治疗,并在先前的全身治疗后或之后疾病进展;以及具有晚期,不可切除或转移性胰腺腺癌的成年人,在先前或之后具有疾病进展的NRG1基因融合

761352s001lbl.pdf - accessdata.fda.gov
警告:胚胎-胎儿毒性 1 适应症和用法 8 特定人群的使用 1.1 晚期不可切除或转移性 NRG1 融合- 8.1 妊娠阳性非小细胞肺癌 8.2 哺乳 1.2 晚期不可切除或转移性 NRG1 融合- 8.3 具有生殖潜力的女性和男性阳性胰腺腺癌 8.4 儿科用途 2 剂量和给药 8.5 老年人使用 2.1 患者选择 11 说明 2.2 开始使用 BIZENGRI 前的建议评估 12 临床药理学 2.3 推荐剂量 12.1 作用机制 2.4 建议的术前用药 12.2 药效学 2.5 不良反应的剂量调整 12.3药代动力学 2.6 制备 12.6 免疫原性 2.7 给药 13 非临床毒理学 3 剂型和强度 13.1 致癌作用、诱变作用、生育能力受损 4 禁忌症 14 临床研究 5 警告和注意事项 14.1 晚期不可切除或转移性 NRG1 融合 5.1 输液相关反应/超敏反应/过敏性阳性非小细胞肺癌反应 14.2 晚期不可切除或转移性 NRG1 融合 5.2 间质性肺病/肺炎阳性胰腺腺癌 5.3 左心室功能障碍 16 如何供应/储存和处理 5.4 胚胎-胎儿毒性 17患者咨询信息 6 不良反应

不同植物中常见的免疫反应节点
二核苷酸糖基水解酶 (NADase) 可产生多种核苷酸衍生的信号分子 ( 5 , 6 )。这些衍生物被进一步加工成短寿命产物,根据其结构,这些产物可作为选择性配体,驱动由脂肪酶样蛋白 EDS1 (增强疾病易感性 1) 和 SAG101 (衰老相关基因 101) 或 PAD4 (植物抗毒素缺乏 4) ( 5 , 6 ) 组成的预先形成的蛋白质异二聚体发生特定重排。然后,两种类型的 EDS1 异二聚体会选择性地募集所谓的“辅助 NLR”,在 EDS1-PAD4 的情况下称为 ADR1(激活抗病性 1),在 EDS1-SAG101 的情况下称为 NRG1(氮必需基因 1)。然后 NRG1 和 ADR1 寡聚化并形成膜定位钙通道,从而激活下游免疫反应,特别是对于 NRG1 而言,导致受感染植物细胞死亡(7,8)。

辅助 NLR 靶向细胞器膜来触发免疫
在植物中,NLR(核苷酸结合域和富含亮氨酸重复序列)蛋白通过形成聚集在质膜上的抗性小体来执行先天免疫。然而,NLR 抗性小体靶向其他细胞膜的程度尚不清楚。在这里,我们表明辅助 NLR NRG1 与多个细胞器膜结合以触发先天免疫。与其他辅助 NLR 相比,NRG1 和密切相关的 RPW8 样 NLR(CC R -NLR)具有延长的 N 端和独特的序列特征,使它们能够组装成比典型的卷曲螺旋 NLR(CC-NLR)抗性小体更长的结构。活化的 NRG1 通过其 N 端 RPW8 样结构域与单膜和双膜细胞器结合。我们的研究结果表明,植物 NLR 抗性小体在各种细胞膜位点聚集以激活免疫。

神经rest和儿子:神经rest细胞和雪旺细胞前体在发育和腺体胚胎发生中的作用
神经rest是一群多能迁移细胞的种群,在神经术期间从神经板的边界分离,并分化成成人生物体中各种器官的细胞(图1;表1)(他,1868年)。根据新的头部假设,正是神经波峰和表皮姿势的外观导致了弦脉的多样化和广泛分布(Gans and Northcutt,1983; Martik and Bronner,2021年)。神经rest细胞经历上皮 - 间质转变的阶段,并开始迁移到身体的远处。NCC与日益增长的神经以及转录特征的变化(SNAI1,SOX9/ 10,FOXD3,PAX3,PAX3和其他NCCS; SOX10,SOX2,SOX2,NRG1)的接触,SOX10,SOX2,NRG1,NRG1,NRG1),结果是Schwann细胞前体的形成,其发展依赖于AXT的迁移,并依赖于AXT的迁移。取决于其起源和定向迁移的位置,整个神经rest细胞(NCC)的种群可以分为几组:颅,树干,心脏和迷走NCCS(Achilleos and Trainor,2012年)。在哺乳动物中,颅NCC会产生颌骨和内耳的软骨和骨结构(Couly等,1998; Freyer等,2011)。此外,该细胞种群产生了牙齿的牙本质,额骨过程的骨骼以及颅神经的周围神经元和神经胶质(Leitevre,1978; Chai et al。,2000;Méndez-Maldez-Maldez-Maldonado et al。,2020)。颅内NCC还分化为The skeletogenic potential of the cranial neural crest has been extensively studied and documented from vertebrates, although cells of the trunk neural crest may also contribute to skeletal components in some animals like the contribution of NCCs to the differentiation of the plastron bones (abdominal carapace bones) of the turtle Trachemys scripta ( Cebra- Thomas et al., 2007 ).

NRG1Fusion的临床病理学特征 - ...
在8148名患者中,在22例患者中鉴定出NRG1融合(0.27%)。患者的平均年龄为59岁(范围为32至78岁),男女比率为1:1.2。肺是最常观察到的主要部位(n = 13),被胰腺片型(n = 3)降低,胃肠道(n = 2,胃和直肠每个),卵巢(n = 2),乳房(n = 1),乳房(n = 1)和软组织(n = 1)。在组织学上,所有肿瘤均表现出腺癌组织学,除了一例肉瘤。CD74(n = 8)和SLC3A2(n = 4)是最常见的融合伙伴。主要的特征包括少于三个同时发生的遗传改变,低肿瘤突变负担和低编程的死亡配体1表达。在NRG1融合患者中观察到各种临床反应。

小家庭,重大影响:RNL助手NLR及其在植物先天免疫中的重要性
通量,活性氧的产生和有丝分裂原激活的蛋白激酶激活[1]。最近的研究表明,2受体系统的相互依赖性和相互增强[2,3]。基于其N末端结构域及其系统发育,NLR在盘绕型圈(CC)结构域,Toll-like/interleukin-1受体耐药性(TIR)结构域中被构成,对白粉病(CC R)的耐药性(CC R)域的耐药性包含NLR,含有NLR,含有AS CNLS,TONLS,the and cnls for and thls for and for and thls from thls&tnls for and。在拟南芥中(以下称为Arabidopsis),多个PRR和效应子传感NLR(某些CNL和所有测试的TNL)需要存在RNL,也称为Helper NLR,以激活全部免疫力[5,6]。rnls形成一个由2个亚家族组成的小而进化保守的进化枝,活化的抗耐药性1(ADR1)和N需求基因1(NRG1)家族,它们在血管植物的发散之前已有分离[4]。拟南芥基因组径流3 ADR1和2 NRG1全长基因需要完全免疫[7-9]。尽管RNL仅代表大多数被子植物中NLR基因库的一小部分[4,10],但对于植物而言,它们至关重要。在这里,我们重点介绍了RNL在免疫过程中的功能以及讨论RNL激活机制的最新发现。