XiaoMi-AI文件搜索系统
World File Search SystemSpike
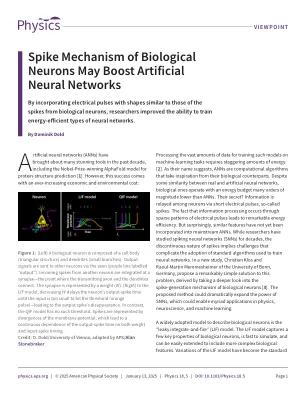
生物神经元的尖峰机制可能会促进人工神经网络
图1 :(左)生物神经元由细胞体(三角形结构)和树突(小分支)组成。输出信号通过轴突(标记为“输出”)发送到其他神经元。来自另一个神经元的传入尖峰集成在突触处,即传输轴突和树突连接的点。突触由重量(W)表示。(右)在LIF模型中,降低W会延迟神经元的输出尖峰时间,直到输入太小而无法达到阈值(橙色脉冲)(橙色脉冲),并导致输出尖峰的消失。相反,QIF模型没有这样的阈值。尖峰由膜电位的差异表示,这导致输出跨度时间对重量和输入跨度的时机的连续依赖性。信用:D。Dold/维也纳大学;由APS/ Alan Stonebraker div>改编

SpikeDeep-classifier:一种基于深度学习的全自动离线脉冲分类算法
摘要 目的。电极设计的进步已导致微电极阵列具有数百个通道,可用于单细胞记录。在由此产生的电生理记录中,每个植入电极可以记录一个或多个神经元的尖峰活动 (SA) 以及背景活动 (BA)。本研究的目的是分离每个神经源的 SA。此过程称为尖峰排序或尖峰分类。高级尖峰排序算法非常耗时,因为在流程的各个阶段都需要人工干预。当前方法缺乏泛化能力,因为超参数的值并不固定,即使对于同一受试者的多个记录会话也是如此。在本研究中,提出了一种称为“SpikeDeep-Classifier”的全自动尖峰排序算法。所有评估数据的超参数值都保持不变。方法。提出的方法基于我们之前的研究 (SpikeDeeptector) 和一种新颖的背景活动拒绝器 (BAR),它们都是监督学习算法和无监督学习算法 (K-means)。 SpikeDeeptector 和 BAR 分别用于提取有意义的通道并从提取的有意义的通道中去除 BA。一旦从数据中完全去除 BA,聚类过程就会变得简单。然后,对仅来自神经源的剩余数据应用具有预定义最大聚类数的 K 均值。最后,使用基于相似性的标准和阈值来保留不同的聚类并合并看起来相似的聚类。所提出的方法称为聚类接受或合并 (CAOM),它只有两个超参数(最大聚类数和相似性阈值),在调整后对于所有评估数据保持不变。主要结果。我们将算法的结果与真实标签进行了比较。该算法在人类患者数据和公开可用的标记非人类灵长类动物 (NHP) 数据集上进行了评估。BAR 在人类患者数据集上的平均准确率为 92.3%,在 (K-means + CAOM) 之后进一步降低到 88.03%。此外,BAR 在公开可用的 NHP 标记数据集上的平均准确率为 95.40%,经过 (K-mean + CAOM) 后降至 86.95%。最后,我们将 SpikeDeep-Classifier 的性能与两位人类专家进行了比较,其中 SpikeDeep-Classifier 产生了可比的结果。意义。SpikeDeep-Classifier 在不同物种、不同大脑区域的多个记录会话的数据集上进行了评估

重新定位靶向刺突糖蛋白的配体作为抗 SARS-CoV-2 的潜在药物
从构象上看,刺突糖蛋白以同源三聚体的形式排列在病毒表面 [29]。当 RBM 被隐藏时,构象称为向下(受体不可接近)(见图 1C)。然而,同源三聚体是不对称的,因为它们不断进行结构重排(向上构象),以将病毒膜与宿主细胞膜融合 [13]。当两个 RBD 结构域被隐藏(受体不可接近)时,一个 RBD 结构域暴露(受体可接近),称为向上构象(见图 1D)。这是因为 S1 的 RBD 经历了铰链状运动 [32]。在 SARS-CoV 中,有两个铰链位点被鉴定(铰链 1 位点(354-361)和铰链 2 位点(552-563),它们负责上下切换

两组分峰纳米粒子疫苗可保护猕猴免受SARS-COV-2感染
两个成分尖峰Nanoopartic疫苗可保护猕猴免受SARS-COV-2感染的菲利森特(J. M. Brucker)1, *, *,Pauline Maisons 2, *, *,Nathale-Nathale-Stryeddreeddre-Bosqut 2,Marous Grobbes 1 Claireaux 1 Claireaux 1,Marlon Yasunori Wasunoraabe 4.5,Gius Sen Strates 1,Cynthia A Schermer 1,Breemen 1,Rashmi Ravichandran 8.9,Tom G. Canis 1,Jellen 1,Niidhal Kahlaouu 2,Kahlaouu 2,Vaehal Contres 2,Vaehal Contres 2,Julien Villaudy 10,julaudy Villaudy 10,kkwenten scopes 1,bert in U. U. Hayman的7,Deric Ginoux 3,Andrew B.Ward 6,Max Crispin 4,Neil P. King 8.9,Sylvie of der 11,Marit J. of Gils 1,Riger Less 1,销售作者。 电子邮件:r.w.have@amsterdamc.nl(R.W.S. ); bearing.le- girn@cea.fr(r.l.g。)Ward 6,Max Crispin 4,Neil P. King 8.9,Sylvie of der 11,Marit J. of Gils 1,Riger Less 1,销售作者。电子邮件:r.w.have@amsterdamc.nl(R.W.S.); bearing.le- girn@cea.fr(r.l.g。)

通用 DNA 适体可有效抑制 Spike 蛋白/hACE2 相互作用
严重急性呼吸综合征冠状病毒 2 (SARS-CoV-2) 使用其刺突蛋白通过人血管紧张素转换酶 2 (hACE2) 附着到宿主细胞上。可以通过设计一种可以阻断刺突蛋白和 hACE2 之间相互作用的抑制剂来阻止病毒感染。如图 1 所示,一个刺突蛋白三聚体包含三个刺突蛋白,每个刺突蛋白由亚基 S1 和 S2 组成。S1 由 S1A 和 S1B 组成(图 1A),其中 S1B 也称为受体结合域 (RBD),与 hACE2 建立直接相互作用。1 此外,S2 亚基在介导病毒膜与宿主细胞融合方面发挥作用。因此,病毒进入是通过一系列事件完成的,即 S1 与 hACE2 结合,然后触发 S2 将其构象改变为更稳定的融合后状态并允许病毒进入宿主细胞。 2–4 由于 S1 直接与 hACE2 相互作用,许多研究小组一直在积极致力于发现各种生物分子,如抗体 5–10 或适体 11–16,以有效阻断 S1 和 hACE2 之间的相互作用。

小麦蔗糖合酶基因tasus1是每个峰值晶粒数的决定因素
植物分子生理学的关键实验室,植物学研究所,中国科学院,北京100093,中国中国国家植物园,北京100093,c国家植物细胞的主要实验室 Sciences, Chinese Academy of Agricultural Sciences (CAAS), Beijing 100081, China e University of Chinese Academy of Sciences, Beijing 100049, China f International Maize and Wheat Improvement Center (CIMMYT) China Office, c/o CAAS, Beijing 100081, China g CAS-JIC Centre of Excellence for Plant and Microbial Science (CEPAMS), Institute of Genetics and Developmental生物学,中国科学院,北京100101,中国植物分子生理学的关键实验室,植物学研究所,中国科学院,北京100093,中国中国国家植物园,北京100093,c国家植物细胞的主要实验室 Sciences, Chinese Academy of Agricultural Sciences (CAAS), Beijing 100081, China e University of Chinese Academy of Sciences, Beijing 100049, China f International Maize and Wheat Improvement Center (CIMMYT) China Office, c/o CAAS, Beijing 100081, China g CAS-JIC Centre of Excellence for Plant and Microbial Science (CEPAMS), Institute of Genetics and Developmental生物学,中国科学院,北京100101,中国

小麦BZIPC1与FT2相互作用,并有助于调节每个峰值的蜘蛛网数
抽象关键信息小麦转录因子BZIPC1与FT2相互作用,并影响Spikelet和每个峰值的晶粒数。我们确定了一个天然等位基因,对这两个经济上重要的特征具有积极影响。在小麦中的基因开花基因座T2(FT2)中的功能丧失突变和自然变异已被证明会影响每个峰值(SNS)的尖峰数。 然而,尽管其他类似FT的小麦蛋白与来自A组的含BZIP的转录因子相互作用,但FT2不与任何一个相互作用。 在这项研究中,我们将酵母2杂交筛选带有FT2作为诱饵,并从C-Group中鉴定出含BzipC1的基于BZIPC1的基因BZIP的转录因子。 在C组中,我们确定了四个进化枝,包括与不同的FT相互作用的小麦蛋白,例如像编码的蛋白一样。 BZIPC1和FT2表达在发育中的峰值中部分重叠,包括花序分生组织。 在BZIPC-A1和BZIPC-B1(BZIPC1)中的功能丧失突变在四倍体小麦中导致SNS的急剧减少,对标题日期的影响有限。 分析BZIPC-B1(TRAESCS5B02G444100)区域的自然变化区域显示,三种主要的单倍型(H1-H3),H1单倍型显示出比H2和H3单倍型的SNS明显更高,每个峰值的晶粒数明显更高,每个峰值的晶粒数明显更高。 H1单倍型的有利作用也得到了其从祖先培养的四倍体到现代四倍体和六比小麦品种的频率增加的支持。在小麦中的基因开花基因座T2(FT2)中的功能丧失突变和自然变异已被证明会影响每个峰值(SNS)的尖峰数。然而,尽管其他类似FT的小麦蛋白与来自A组的含BZIP的转录因子相互作用,但FT2不与任何一个相互作用。在这项研究中,我们将酵母2杂交筛选带有FT2作为诱饵,并从C-Group中鉴定出含BzipC1的基于BZIPC1的基因BZIP的转录因子。在C组中,我们确定了四个进化枝,包括与不同的FT相互作用的小麦蛋白,例如像编码的蛋白一样。BZIPC1和FT2表达在发育中的峰值中部分重叠,包括花序分生组织。在BZIPC-A1和BZIPC-B1(BZIPC1)中的功能丧失突变在四倍体小麦中导致SNS的急剧减少,对标题日期的影响有限。分析BZIPC-B1(TRAESCS5B02G444100)区域的自然变化区域显示,三种主要的单倍型(H1-H3),H1单倍型显示出比H2和H3单倍型的SNS明显更高,每个峰值的晶粒数明显更高,每个峰值的晶粒数明显更高。H1单倍型的有利作用也得到了其从祖先培养的四倍体到现代四倍体和六比小麦品种的频率增加的支持。我们开发了两个非同义SNP的标记,这些标记将H1单倍型中的BZIPC-B1B等位基因与所有其他单倍型中存在的祖先BZIPC-B1A等位基因区分开。这些诊断标记是加速在面食和面包小麦育种计划中的有利BZIPC-B1B等位基因部署的有用工具。

大脑刺激与持续的振荡竞争,以控制灵长类动物大脑中的尖峰时序
经颅交流电流刺激(TAC)通常用于增强脑节律,以期改善行为性能。不幸的是,这些干预措施通常会产生高度可变的结果。在这里,我们通过在警报非人类灵长类动物中记录单个神经元来确定这种变异性的关键来源。我们发现,TACS似乎与大脑的内源性振荡竞争以控制尖峰时序,而不是增强节奏活动。具体而言,当刺激的强度相对于内源性振荡较弱时,TACS实际上会降低尖峰的节奏性。但是,当刺激相对较强时,TACS对尖峰活动施加了自身的节奏。因此,TAC的作用明确地取决于神经夹带的强度,内源性振荡在行为状态和大脑区域之间差异很大。未经仔细考虑这些因素,试图将外部节奏施加到特定的大脑区域,通常可能与预期效应相反。

脑刺激与持续振荡竞争,以控制灵长类动物大脑中的脉冲时间
经颅交流电刺激 (tACS) 是一种常用的非侵入性脑活动调节方法。具体来说,tACS 经常被用作一种有针对性的干预手段,通过增强特定频率的神经振荡来影响特定行为。然而,这些干预手段往往产生高度可变的结果。在这里,我们为这种可变性提供了一个可能的解释:tACS 与大脑的持续振荡相竞争。利用来自警觉的非人类灵长类动物的神经记录,我们发现,当神经放电独立于持续的脑振荡时,tACS 很容易同步脉冲活动,但当神经元强烈同步于持续的振荡时,tACS 通常会导致同步减少。因此,即使刺激方案是固定的,tACS 也可以对神经活动产生截然不同的结果。数学分析表明,这种竞争很可能在许多实验条件下发生。因此,试图将外部节奏强加于大脑往往会产生完全相反的效果。

一种用于实现随机二进制尖峰时间相关可塑性的 CMOS-忆阻器混合系统
本文介绍了一个完全实验性的混合系统,其中使用定制的高阻态忆阻器和采用 180 nm CMOS 技术制造的模拟 CMOS 神经元组装了一个 4 × 4 忆阻交叉脉冲神经网络 (SNN)。定制忆阻器使用 NMOS 选择晶体管,该晶体管位于第二个 180 nm CMOS 芯片上。一个缺点是忆阻器的工作电流在微安范围内,而模拟 CMOS 神经元可能需要的工作电流在皮安范围内。一种可能的解决方案是使用紧凑电路将忆阻器域电流缩小到模拟 CMOS 神经元域电流至少 5-6 个数量级。在这里,我们建议使用基于 MOS 阶梯的片上紧凑电流分配器电路,将电流大幅衰减 5 个数量级以上。每个神经元之前都添加了这个电路。本文介绍了使用 4 × 4 1T1R 突触交叉开关和四个突触后 CMOS 电路的 SNN 电路的正确实验操作,每个电路都有一个 5 个十进制电流衰减器和一个积分激发神经元。它还演示了使用此小型系统进行的一次性赢家通吃训练和随机二进制脉冲时间依赖可塑性学习。