XiaoMi-AI文件搜索系统
World File Search SystemTestis
果蝇睾丸生态裂干细胞
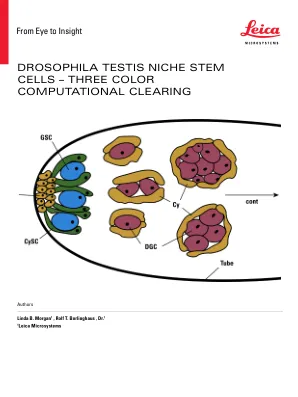
图2:果蝇睾丸中的干细胞生态位。褐色:轮毂细胞HC。蓝色:种系干细胞GSC。绿色:囊肿干细胞CYSC。红色:区分生殖细胞。棕色:囊肿。利基位于睾丸管的远端。种系干细胞和囊肿干细胞已连接并增殖,并由几个组成“轮毂”的细胞锚定。区分时,一组一组分化的种系细胞与两个囊肿细胞形成复合物。囊肿充当区分生殖细胞的容器,该容器经历了细胞分裂,构成了囊肿中包含的精子祖细胞的包装。这种发展由持续细胞分裂的箭头指示(续)。

pd- 1通过IFN-γ-stat1- irf1- lpcat3
抽象滑膜肉瘤(SS)是一种罕见的癌症,对儿童和年轻人的影响不成比例。癌睾丸抗原(CTA)是在胚胎发育早期表达的蛋白质,但通常在正常组织中表达。它们在许多不同的癌症类型中异常表达,并且是免疫疗法的有吸引力的治疗靶标。CTA在SS中以高水平表示。这种高水平的CTA表达使SS成为旨在利用免疫系统识别异常CTA表达并与癌症作斗争的治疗策略的理想癌症。关键临床试验现在正在进行中,从当前的护理标准中大大改变了SS管理和治疗的景观。在这篇综述中,我们描述了针对SS中CTA的基本原理,重点是NY-ESO-1和MAGE-A4,即疫苗和基于T细胞受体的疗法的现状,并考虑为未来开发的新兴机会。

黑色素瘤和肺癌中的睾丸抗原表达
摘要癌症/睾丸抗原(CTA)在黑色素瘤和肺癌中广泛表达,成为这些恶性肿瘤中疫苗接种策略和基于T细胞的疗法的有希望的靶标。尽管认识到肿瘤内异质性对免疫疗法的临床反应的基本影响,但我们对CTA表达中肿瘤内异质性的理解仍然有限。我们采用单细胞mRNA测序来描述临床衍生的黑色素瘤和肺癌样品中癌细胞的CTA表达谱。我们的发现显示了CTA表达中高度的肿瘤内转录异质性。在黑色素瘤中,每个细胞都表达至少一个CTA。 但是,大多数单独的CTA,包括广泛使用的治疗靶标NY-ESO-1和MAGE,都局限于细胞的亚群,并在其表达中不协调,从而导致具有不同CTA谱的癌细胞的镶嵌物。 但是,观察到协调的表达主要在高度结构和进化相关的CTA基因中。 重要的是,在黑色素瘤中均匀地表达了CTA的较小子集,包括Prame和Gage和Mage-A家族的几个成员,强调了它们作为治疗靶标的潜力。 在肺癌中还观察到了CTA表达的广泛异质性。 然而,CTA阳性癌细胞的频率明显较低,并且仅在该癌症类型的五个肿瘤之一中鉴定出同质表达的CTA。在黑色素瘤中,每个细胞都表达至少一个CTA。但是,大多数单独的CTA,包括广泛使用的治疗靶标NY-ESO-1和MAGE,都局限于细胞的亚群,并在其表达中不协调,从而导致具有不同CTA谱的癌细胞的镶嵌物。协调的表达主要在高度结构和进化相关的CTA基因中。重要的是,在黑色素瘤中均匀地表达了CTA的较小子集,包括Prame和Gage和Mage-A家族的几个成员,强调了它们作为治疗靶标的潜力。在肺癌中还观察到了CTA表达的广泛异质性。然而,CTA阳性癌细胞的频率明显较低,并且仅在该癌症类型的五个肿瘤之一中鉴定出同质表达的CTA。我们的发现强调了在免疫疗法开发和临床测试中仔细选择CTA目标选择的必要性,并提供了一个合理的框架来识别最有希望的候选人。

基于 CRISPR/Cas9 的小鼠基因组编辑揭示了 13 个睾丸
1 日本大阪大学微生物疾病研究所实验基因组研究系,2 日本大阪大学医学研究生院,3 美国德克萨斯州休斯顿贝勒医学院药物发现中心,4 美国德克萨斯州休斯顿贝勒医学院病理学和免疫学系,5 日本大阪大学药学研究生院,6 日本名古屋名古屋市立大学医学研究生院比较与实验医学系,7 日本茨城县筑波市筑波大学医学院解剖学与胚胎学系,8 美国德克萨斯州休斯顿休斯顿大学克利尔莱克分校生物与生物技术系,9 日本东京大学医学科学研究所

果蝇睾丸中Mitf/TFEB的减数分裂活性随年龄增长而下降
诱导 GFP 表达。C、D. 成年年轻果蝇睾丸中的 DNA(Hoechst)和 4Mbox-GFP(Mitf 活性报告基因)的代表性图像。D 中勾勒出了 C 中精原细胞和精母细胞的放大区域。C 中勾勒出了囊细胞核。DE 中的虚线标出了有丝分裂到减数分裂的转变。对成年年轻雄性有丝分裂精原细胞(n = 10 个睾丸中的 50 个精原细胞)和减数分裂精母细胞(n = 10 个睾丸中的 50 个精母细胞)中 4MBox-GFP 强度的量化。平均值 ± SD p < 0.0001,Mann-Whitney U 检验。 F. 量化年轻男性(n =60 个精母细胞,来自 12 个睾丸)和老年男性(n =80 个精母细胞,来自 16 个睾丸)精母细胞中 4Mbox-GFP 强度。平均值 ± SD p < 0.0001,Welch t 检验。G. 年轻和老年男性精母细胞中 DNA(Hoechst)和 4Mbox-GFP(Mitf 活性报告基因)的代表性图像。H. 量化年轻男性(n =60 个精母细胞,来自 12 个睾丸)和老年男性(n =65 个精母细胞,来自 13 个睾丸)精母细胞中 VhaSFD-GFP 强度。平均值 ± SD p < 0.0001,Welch t 检验。I. 年轻和老年男性精母细胞中 DNA(Hoechst)和 VhaSFD-GFP 的代表性图像。条,20 µm。

小鼠围产期睾丸发育中的单细胞染色质可及性景观
1 香港中文大学医学院生物医学学院发育及再生生物学项目,香港沙田,香港;2 福建医科大学医学科技与工程学院,福建,中国;3 香港中文大学医学院生物医学学院癌症生物学及实验治疗学项目,香港沙田,中国;4 广州生物医药与健康研究院广州再生医学与健康生物园实验室,广州,中国;5 哈德逊医学研究所生殖健康中心生殖干细胞生物学实验室,墨尔本,澳大利亚;6 香港中文大学化学病理学系,香港新界沙田,中国

KU70,SIRT1和SIRT6蛋白在糖尿病大鼠睾丸组织中的作用 技术对食物浪费的影响:智能包装 ortaokulÖğrencilininetkinvatandaşlıkKavramına所以 do钠葡萄糖共转运蛋白-2(SGLT-2)抑制剂影响... Erdoğan,S。和M. Y.(2024)。 bitkiselkaynaklılifler ve胃肠道西姆·üzerineetkileri。 BütünleyiciveanadoluTıbbıDergisi,5(SbüH 全面质量管理实践在... 上的重要性
摘要:糖尿病男性不孕/不育是糖尿病的重要并发症。尚未对负责这种并发症的分子机制进行彻底研究。我们旨在阐明KU70,SIRT1和SIRT6蛋白在糖尿病睾丸中的作用。Sprague -Dawley雄性大鼠在稳定的实验室条件下保持。将大鼠分为对照组(n = 8)和一个糖尿病组(n = 8,单剂量为50 mg/kg链霉菌素)。在1个月的实验期结束时,在麻醉下处死动物。将两个睾丸均已去除,轻轻处理,并通过电子传输显微镜和蛋白质印迹研究。血液样本进行生化分析。组织病理学分析表明,在糖尿病组中,diaphragmatic小管直径和血清睾丸激素水平降低。ku70免疫反应性在统计学上显着增加,而与对照组观察到的相比,SIRT1和SIRT6表达显着降低。这是首次检查KU70,SIRT1和SIRT6在糖尿病睾丸组织中的表达的第一项研究。根据结果,KU70,SIRT1和SIRT6可能在糖尿病睾丸组织中细胞凋亡中起重要作用。在其他定量研究中应进一步研究这些蛋白质的重要性。

癌症/睾丸抗原 55 是癌细胞增殖和线粒体 DNA 维持所必需的
癌症/睾丸抗原 (CTA) 代表一组蛋白质,其在生理条件下的表达仅限于睾丸,但在许多人类癌症中被激活。此外,据观察,多个 CTA 的共同表达会使患者的预后恶化。据报道,有五种 CTA 作用于线粒体,我们最近报道了 67 种 CTA 编码的 147 个转录本,这些转录本编码了可能针对线粒体的蛋白质。其中,我们确定了 CT55 编码的两种异构体,其功能尚不明确。首先,我们发现表达野生型 CT55 的肿瘤患者生存率较低。此外,CT55 沉默会显著降低细胞增殖。其次,为了研究 CT55 对线粒体的作用,我们首先表明,由于存在不明确的 N 端靶向信号,CT55 定位于线粒体和内质网 (ER)。然后,我们表明 CT55 沉默会降低 mtDNA 拷贝数并延迟急性耗竭后的 mtDNA 恢复。此外,CT55 启动子的去甲基化会增加其表达,进而增加 mtDNA 拷贝数。最后,我们测量了 NCI-60 细胞系中的 mtDNA 拷贝数,并筛选出表达与 mtDNA 量密切相关的基因。我们将 CT55 确定为第二高的相关性命中。此外,我们还表明,与 siRNA 乱序对照 (siCtrl) 治疗相比,CT55 特异性 siRNA (siCT55) 治疗下调了

除草剂利谷隆对热带爪蟾大脑和睾丸 DNA 甲基化谱的雄性传递跨代效应
除草剂利谷隆可对非洲爪蟾(Xenopus Tropicalis)产生内分泌干扰作用,包括从未接触过该污染物的后代。这些影响跨代传递的机制有待进一步研究。在这里,我们研究了大脑和睾丸 DNA 甲基化谱的跨代改变,这些改变是从发育过程中接触到环境相关浓度利谷隆的祖父那里遗传下来的。简化代表性亚硫酸氢盐测序 (RRBS) 揭示了成年雄性 F2 代大脑 (3060 个 DMR) 和睾丸 (2551 个 DMR) 中的许多差异甲基化区域 (DMR)。大脑中参与生长激素 ( igfbp4 ) 和促甲状腺激素信号传导 ( dio1 和 tg ) 的关键基因存在差异甲基化,并与体型、体重、后肢长度和血糖水平的表型改变相关,表明这些甲基化变化可能是利谷隆跨代效应的潜在介质。睾丸 DMR 存在于精子发生、减数分裂和生殖细胞发育所必需的基因( piwil1 、 spo11 和 tdrd9 )中,其甲基化水平与每个曲细精管的生殖细胞巢数量相关,这是精子发生中断的终点。DMR 还存在于调节表观遗传景观的机制(包括 DNA 甲基化)的几个基因中

N6-甲基读丹代氨酸读取器IGF2BP2修饰HMMR ...
睾丸负责精子产生和雄激素合成。睾丸发育和功能的异常导致性发展和男性不育症的疾病。当前,没有用于对睾丸进行建模的体外系统。在这里,我们使用Transwell插入物从新生小鼠初级睾丸细胞中产生睾丸类器官,并表明这些类型器可以生成类似小管的结构和类似于体内睾丸的细胞组织。基因表达分析表明了一种概括体内睾丸中观察到的特征。胚胎睾丸细胞,但没有成年睾丸细胞也能够形成器官。这些类器官可以在培养物中维持8-9周,并显示出进入减数分裂的迹象。我们进一步开发了定义的培养基组成,这些培养基组成促进了未成熟的Sertoli细胞和Leydig细胞状态,从而在体外实现了器官成熟。这些睾丸类器官是一种有前途的模型系统,用于睾丸发育和功能的基础研究,并在阐明和治疗发育性疾病和不育的情况下进行了翻译应用。