XiaoMi-AI文件搜索系统
World File Search SystemWx
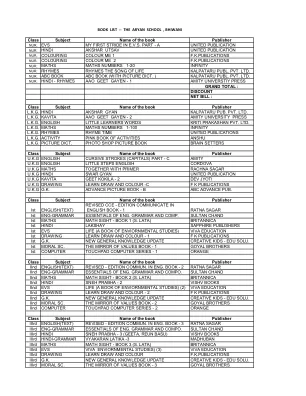
书籍出版商的类主题名称
书籍的班级名称出版商vii the English(文本)Honey Comb N.C.E.R.T班级-7英语A外星人手N.C.E.R.T班级-8 VII TH ENG -GRAMMAR ENG的ENGER ESTENCES。语法和综合。SULTAN CHAND VII th HINDI BASANT - PART - 2 N.C.E.R.T CLASS - 7 VII th HINDI-GRAMMAR VYAKARAN LATIKA - 7 MADHUBAN VII th MATHS TEXT BOOK FOR CLASS - VII N.C.E.R.T CLASS - 7 VII th SCIENCE TEXT BOOK FOR CLASS - VII N.C.E.R.T CLASS - 7 VII th SCIENCE WX LAB MANUAL科学-7 Saraswati VII TH G.K.新的常识更新创意儿童-Edu Solu。vii The Computer TouchPad计算机系列-7 Orange VII TH S.St.社会和政治生活-II N.C.E.R.T班-7 VII TH S.St.我们的环境(GEO)N.C.E.R.T类-7 VII TH S.St.我们的过去-2 N.C.E.R.T类-7 VII TH Sanskrit Deep Manika Part -2 Saraswati

Squamish遗产管理策略2023
2.5提供有关考古遗产资源管理的公共教育,意识和获得发展批准信息。该地区有责任向公众教育有关SḵWX̱Wú7 -Mesh遗产,并防止对可能的未知和未发现的考古遗址避免影响。2.6为该地区进行考古概述评估(AOA),AOA汇编了有关记录的考古遗址位置的现有知识,历史原住民的土地使用以及文化和环境考虑。考古概述和影响评估的目的是确保第一民族的文化遗产资源得到保存并减轻发展影响。2.7为私有财产所有者提供信息,该地区可以通过在准备开发计划时为他们提供相关信息,从而为开发商,业主和设计专业人士提供遗产保护。1.4更新遗产管理策略2.8创建邻里历史上下文陈述应定期审查和更新遗产管理策略,以确保它具有最新的最佳实践和现场的新思维。

使用皮秒 X 射线可视化层状材料中埋藏界面的能量转移
了解纳米级热传播的基本原理对于下一代电子产品至关重要。例如,已知层状材料的弱范德华键会限制其热边界导率 (TBC),从而成为散热瓶颈。本文提出了一种新的非破坏性方法,使用时间分辨的光致热应变 X 射线测量来探测纳米级晶体材料中的热传输。该技术通过测量光激发后 c 轴晶格间距的变化,直接监测晶体中随时间的温度变化以及随后跨埋层界面的弛豫。研究了五种不同的层状过渡金属二硫属化物 MoX 2 [X = S、Se 和 Te] 和 WX 2 [X = S 和 Se] 的薄膜以及石墨和 W 掺杂的 MoTe 2 合金。在室温下,在 c 平面蓝宝石衬底上发现 TBC 值在 10–30 MW m − 2 K − 1 范围内。结合分子动力学模拟,结果表明高热阻是界面范德华键合较弱和声子辐射度较低造成的。这项研究为更好地理解新兴 3D 异质集成技术中的热瓶颈问题奠定了基础。

使用近似刺激呈现技术的稳态视觉诱发电势...
İfĉāmicāë0ë0údk3m7m7mbgn <3Mbgn <3Mbgn <3Mbgn <3Mbgn <3mbgn = 〜3m = 〜3℺«c«c«c«c«c«c«c«c。 Åħ}。 ib¾hs² -2ij2 - #ijzë -ę[0平均ģ+。 u#nd $ 1avos_tvoīð²±ijzó¾hõ¾hõ«0la£out- O - *tr 2 tr 2 tr 2 tr 2! \äę[0úmñ«â€™tâzë -tâ\ \ääääääääääisúëúëúëâ#j#ândzá - *,Øsâo n o n o n onij ssvep \ä0č»ù¼ijċčmñmï-\ässvep \äOiijzë--\ä /¾hâtome -į 1/4ij½½的3¾hs。

飞机规格 - Eagle Aviation
新机使用时间 1564 小时 1564 小时 TBO 3600 小时 3600 小时 道具:(4 叶片 McCauley)新机使用时间 1564 小时 1564 小时 大修使用时间 312 小时 312 小时 外观:整体马特洪峰白色,带城堡银色和黄昏灰色条纹。2010 年。内饰:四个俱乐部座椅,配有安全带便盆,采用 Mammut Bone 皮革制成,机组座椅配有 Aerolamb Camel 羊皮嵌件,Honeycomb II Quarry 地毯,下部侧板采用 Mannut Bone 皮革,装饰条和扶手采用 Eurosoft Charcoal,车顶内衬采用 Ultraleather Milkweed,窗户面板采用 Buffalino 3015 Surfside,窗帘隔断采用 Line Basket Black Pearl。 2010. 航空电子设备: 配备 GAD43E(ADF/DME)和合成视觉的 Garmin G600 PFD Garmin GTN 750 触摸屏 GPS Garmin GNS 530 GPS 导航/通信 TAWS Garmin GDL88 ADS-B 输出 Garmin TAS800 交通系统 Garmin GMA-340 音频面板 升级版 Blackhawk 数字扭矩表 Shadin 数字燃油流量计 Garmin GTX33 E/S 和 GTX330 ES 应答器 ADS-B 输入 GDL69 XM WX 和娱乐 ARC 1000A 自动驾驶仪 Collins WXR-200A 彩色雷达 BF Goodrich WX-1000E Stormscope 选项: 速度堆栈 406 ELT 工厂空调 新型飞行员侧电动挡风玻璃

防寒保暖援助计划政策和程序......
第 1 节:目的和范围 防寒保暖援助计划 (Wx) 的目的是提高低收入人群拥有或居住的单位的能源效率,或为这些人提供可再生能源系统或技术,减少他们的总住宅支出,并改善他们的健康和安全,特别是那些特别脆弱的低收入人群,例如老年人、残疾人、有孩子的家庭、高住宅能源用户以及高能源负担的家庭。10 CFR 440.1。所有联邦计划法规和指导均可在 SCEP 的防寒保暖援助计划中找到。如果印第安纳州专门发布了计划指南,则可以在 MyIHCDA 的防寒保暖中找到这些通知。所有防寒保暖工作都必须遵守能源部的规章制度,除非政策和程序手册、拨款协议和/或计划指南中明确规定了例外情况。任何偏离 DOE 指南、印第安纳州政策和程序手册或印第安纳州防寒保暖现场指南 SWS 对齐版的行为都必须在单位工作前获得 IHCDA 的书面批准。此批准仅适用于提出请求的单位,并且必须放入客户文件中。 第 2 节:奖励 2.1 分配 在绩效期开始时,根据 IHCDA 的分配表向子受助人分配联邦资金。分配表基于子受助人服务区域的人口因素,并确定机构将收到的资金数额。子受助人可根据要求获得额外资金或单独分配。 2.2 绩效期 绩效期是给予子受助人履行所有合同义务的时间。绩效期内有不同的时期,如下所示:
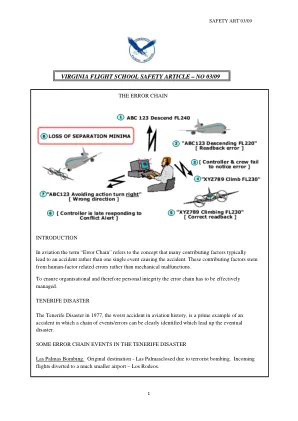
错误链 - 弗吉尼亚飞行学校
加油。荷航 747 决定在滑行道上的 Los Rodeo 加油。与此同时,拉斯帕尔马斯机场已重新开放。加油阻塞了滑行道,使飞机无法起飞,从而导致拥堵,其他机组人员感到沮丧。滑行。由于滑行道上挤满了飞机,荷航和泛美航空不得不在跑道上向后滑行到起飞阈值,并在阈值处转 180 度。对于 747 来说,在 45 米宽的跑道上行驶非常困难。泛美航空跟随荷航沿跑道行驶。他们被指示在滑行道 3 号出口离开跑道。没有标记指示滑行道出口。出口 3 需要 145 度“向后”转弯,让泛美航空回到滑行道上等待起飞的飞机队列中。对于 747 来说,向 3 号出口转 145 度几乎是不可能完成的。天气。在两架 747 滑行过程中,由于低云,WX 恶化。报告的最大水平可视范围为 300 米。通信。塔台发出的 ATC 许可包括“起飞”一词。副驾驶复读许可并通知塔台 - “我们正在起飞”,这意味着他们已准备好起飞。塔台回应“OK”。荷航机长将此解释为继续起飞的许可并打开油门。塔台说“准备起飞 - 我会打电话给你”。此时,泛美航空意识到危险,向塔台传递信息,他们仍在跑道上滑行,阻挡了塔台呼叫荷航准备起飞许可。荷航飞行工程师

2022-2023 年青少年 DTaP-IPV 和 MMR 疫苗接种情况...
S CHO4 KY 克里 1884 1650 87.6 1884 1657 88.0 S CHO4 NC 北科克 1337 1268 94.8 1337 1267 94.8 S CHO4 NSL 北李郡-科克 2097 1975 94.2 2097 1966 93.8 S CHO4 NSL 南李郡-科克 2917 2769 94.9 2917 2753 94.4 S CHO4 WC 西科克 789 709 89.9 789 709 89.9 SE CHO5 CW/KK 卡洛/基尔肯尼 1800 1724 95.8 1800 1721 95.6 SE CHO5 TS蒂珀雷里南部 1268 1098 86.6 1268 1101 86.8 SE CHO5 WD 沃特福德 1694 1479 87.3 1694 1456 86.0 SE CHO5 WX 韦克斯福德 2058 1922 93.4 2058 1926 93.6 E CHO6 CCA1 都柏林南部 1767 1645 93.1 1767 1643 93.0 E CHO6 CCA2 都柏林东南 1715 1691 98.6 1715 1691 98.6 E CHO6 CCA10 威克洛 1693 1592 94.0 1693 1591 94.0 E CHO7 CCA3 都柏林南部城市 1769 1637 92.5 1769 1622 91.7 E CHO7 CCA4 都柏林西南 1766 1460 82.7 1766 1458 82.6 E CHO7 CCA5 都柏林西 2047 1715 83.8 2047 1706 83.3 E CHO7 CCA9 基尔代尔/西威克洛 3738 3512 94.0 3738 3510 93.9 M CHO8 LS/OY Laois/Offaly 2346 2142 91.3 2346 2148 91.6 M CHO8 LD/WH 朗福德/韦斯特米斯 1980 1803 91.1 1980 1803 91.1 NE CHO8 LH 劳斯郡 1837 1638 89.2 1837 1628 88.6 NE CHO8 MH 米斯郡 3036 2768 91.2 3036 2735 90.1 E CHO9 CCA8 都柏林北部 3794 3203 84.4 3794 3187 84.0 E CHO9 CCA7 都柏林中北部 1338 995 74.4 1338 987 73.8 E CHO9 CCA6 都柏林西北部 2889 2250 77.9 2889 2247 77.8

CRISPR/Cas基因编辑技术治疗人类遗传性疾病的临床 ...
[1] Egger G,Liang G,Aparicio A等。人类疾病的表观遗传学和表观遗传疗法的前景。 自然,2004,429:457-63 [2] Varmus H.为基于基因的药物做好准备。 New Engl J Med,2002,347:1526-7 [3] Pogue RE,Cavalcanti DP,Shanker S等。 罕见的遗传疾病:诊断,治疗和在线资源的更新。 Div> Discov今天,2018年,23:187-95 [4] Fischer A,Cavazzana-Calvo M.遗传疾病的基因治疗。 Lancet,2008,371:2044-7 [5] Porteus M.基因组编辑:一种新的人类治疗方法。 Annu Rev Pharmacol Toxicol,2016,56:163-90 [6] Cox DBT,Platt RJ,ZhangF。治疗基因组编辑:前景和挑战。 nat Med,2015,21:121-31 [7] Barrangou R,Fremaux C,Deveau H等。 crispr提供了对原核生物中病毒的抗药性。 Science,2007,315:1709-12 [8] Deltcheva E,Chylinski K,Sharma CM等。 CRISPR RNA通过反式编码的小RNA和宿主因子RNase III成熟。 自然,2011,471:602-7 [9] Cong L,Ran FA,Cox D等。 使用CRISPR/CAS系统的多重基因组工程。 Science,2013,339:819-23 [10] Jinek M,Chylinski K,Fonfara I等。 适应性细菌免疫中可编程的双RNA引导的DNA内切酶。 Science,2012,337:816-21 [11] Maruyama T,Dougan SK,Truttmann MC等。 通过抑制非同源末端连接来提高精确基因组编辑的效率。 nat Biotechnol,2015,33:538-42 [12] Shmakov S,Smargon A,Scott D等。 快照:2类CRISPR-CAS系统。人类疾病的表观遗传学和表观遗传疗法的前景。自然,2004,429:457-63 [2] Varmus H.为基于基因的药物做好准备。New Engl J Med,2002,347:1526-7 [3] Pogue RE,Cavalcanti DP,Shanker S等。罕见的遗传疾病:诊断,治疗和在线资源的更新。Div> Discov今天,2018年,23:187-95 [4] Fischer A,Cavazzana-Calvo M.遗传疾病的基因治疗。Lancet,2008,371:2044-7 [5] Porteus M.基因组编辑:一种新的人类治疗方法。Annu Rev Pharmacol Toxicol,2016,56:163-90 [6] Cox DBT,Platt RJ,ZhangF。治疗基因组编辑:前景和挑战。nat Med,2015,21:121-31 [7] Barrangou R,Fremaux C,Deveau H等。crispr提供了对原核生物中病毒的抗药性。Science,2007,315:1709-12 [8] Deltcheva E,Chylinski K,Sharma CM等。CRISPR RNA通过反式编码的小RNA和宿主因子RNase III成熟。自然,2011,471:602-7 [9] Cong L,Ran FA,Cox D等。使用CRISPR/CAS系统的多重基因组工程。Science,2013,339:819-23 [10] Jinek M,Chylinski K,Fonfara I等。适应性细菌免疫中可编程的双RNA引导的DNA内切酶。Science,2012,337:816-21 [11] Maruyama T,Dougan SK,Truttmann MC等。通过抑制非同源末端连接来提高精确基因组编辑的效率。nat Biotechnol,2015,33:538-42 [12] Shmakov S,Smargon A,Scott D等。快照:2类CRISPR-CAS系统。2类CRISPR-CAS系统的多样性和演变。Nat Rev Microbiol,2017,15:169-82 [13] Makarova KS,Zhang F,Koonin EV。Cell,2017,168:328-328.e1 [14] Zetsche B,Gootenberg JS,Abudayyeh Oo等。CPF1是2类CRISPR- CAS系统的单个RNA引导的内切酶。 Cell,2015,163:759-71 [15] Ran Fa,Cong L,Yan WX等。 使用金黄色葡萄球菌Cas9的体内基因组编辑。 自然,2015,520:186-91 [16] Kim E,Koo T,Park SW等。 在体内基因组编辑中,带有弯曲杆菌的小Cas9直系同源CPF1是2类CRISPR- CAS系统的单个RNA引导的内切酶。Cell,2015,163:759-71 [15] Ran Fa,Cong L,Yan WX等。使用金黄色葡萄球菌Cas9的体内基因组编辑。 自然,2015,520:186-91 [16] Kim E,Koo T,Park SW等。 在体内基因组编辑中,带有弯曲杆菌的小Cas9直系同源使用金黄色葡萄球菌Cas9的体内基因组编辑。自然,2015,520:186-91 [16] Kim E,Koo T,Park SW等。在体内基因组编辑中,带有弯曲杆菌的小Cas9直系同源在体内基因组编辑中,带有弯曲杆菌的小Cas9直系同源

基于CRISPR/Cas9 的激活系统研究进展
816–821。[11] Lowder L,Malzahn A,Qi YP. Rapid evolution of manifold CRISPR systems for plant gene editing. Front Plant Sci, 2016, 7: 1683。[12] Maeder ML,Linder SJ,Cascio VM,等。CRISPR RNA引导的内源性人类基因激活。Nat Methods, 2013, 10(10): 977-979。[13] Lindhout BI,Pinas JE,Hooykaas PJJ,等。利用锌指人工转录因子文库筛选拟南芥同源重组突变体。Plant J, 2006, 48(3): 475-483。[14] Liu WS,Rudis MR,Peng YH,等。合成TAL效应物用于靶向增强植物转基因表达。Plant Biotechnol J,2014,12(4):436-446。[15] Qi LS、Larson MH、Gilbert LA等。将CRISPR重新用作RNA引导平台,用于序列特异性控制基因表达。Cell,2013,152(5):1173-1183。[16] Chylinski K、Le Rhun A、Charpentier E。II型CRISPR-Cas免疫系统的tracrRNA和Cas9家族。RNA Biol,2013,10(5):726-737。[17] Nishimasu H、Cong L、Yan WX等。金黄色葡萄球菌Cas9的晶体结构。Cell,2015,162(5):1113-1126。 [18] Jinek M, Jiang FG, Taylor DW 等. Structures of Cas9 endonucleases reveal RNA-mediated configuration activity. Science, 2014, 343(6176): 1247997。[19] Anders C, Niewoehner O, Duerst A 等. Structural basis of PAM-dependent target DNA identification by the Cas9 endonuclease. Nature, 2014, 513(7519): 569-573。[20] Wang Y, Zhang ZT, Seo SO 等. Gene transcription repression in Clostridium beijerinckii using CRISPR-dCas9. Biotechnol Bioeng, 2016, 113(12): 2739-2743。[21] Bikard D, Jiang WY, Samai P 等.利用工程化的 CRISPR-Cas 系统可编程地抑制和激活细菌基因表达。Nucleic Acids Res,2013,41(15):7429-7437。[22] Didovyk A、Borek B、Tsimring L 等人。利用 CRISPR-Cas9 进行转录调控:原理、进展和应用。Curr Opin Biotechnol,2016,40:177-184。[23] Li ZX、Xiong XY、Li JF。工作死物:将失活的 CRISPR 相关核酸酶重新用作植物中的可编程转录调节剂。aBIOTECH,2020,1(1):32-40。[24] Piatek A、Ali Z、Baazim H 等人。RNA 引导的