XiaoMi-AI文件搜索系统
World File Search Systemcapsid

干扰素诱导的GTPase MXB促进疱疹病毒的capsID拆卸和基因组释放
1德国汉诺威汉诺威医学院病毒学研究所; 2抵抗 - 卓越群,德国汉诺威汉诺威医学院; 3德国慕尼黑慕尼黑技术大学病毒学研究所; 4德国弗莱堡大学弗莱堡大学医学中心病毒学研究所; 5美国普林斯顿大学普林斯顿大学分子生物学系; 6 MRC人类免疫学部门,MRC Weatherall分子医学研究所,Radcliffe医学系,牛津大学,牛津大学,英国; 7德国汉诺威 - 布劳恩斯乔格合作伙伴网站德国感染研究中心(DZIF),德国汉诺威; 8德国汉诺威汉诺威医学院的研究核心单位激光显微镜; 9德国慕尼黑的德国感染研究中心(DZIF),德国慕尼黑1德国汉诺威汉诺威医学院病毒学研究所; 2抵抗 - 卓越群,德国汉诺威汉诺威医学院; 3德国慕尼黑慕尼黑技术大学病毒学研究所; 4德国弗莱堡大学弗莱堡大学医学中心病毒学研究所; 5美国普林斯顿大学普林斯顿大学分子生物学系; 6 MRC人类免疫学部门,MRC Weatherall分子医学研究所,Radcliffe医学系,牛津大学,牛津大学,英国; 7德国汉诺威 - 布劳恩斯乔格合作伙伴网站德国感染研究中心(DZIF),德国汉诺威; 8德国汉诺威汉诺威医学院的研究核心单位激光显微镜; 9德国慕尼黑的德国感染研究中心(DZIF),德国慕尼黑
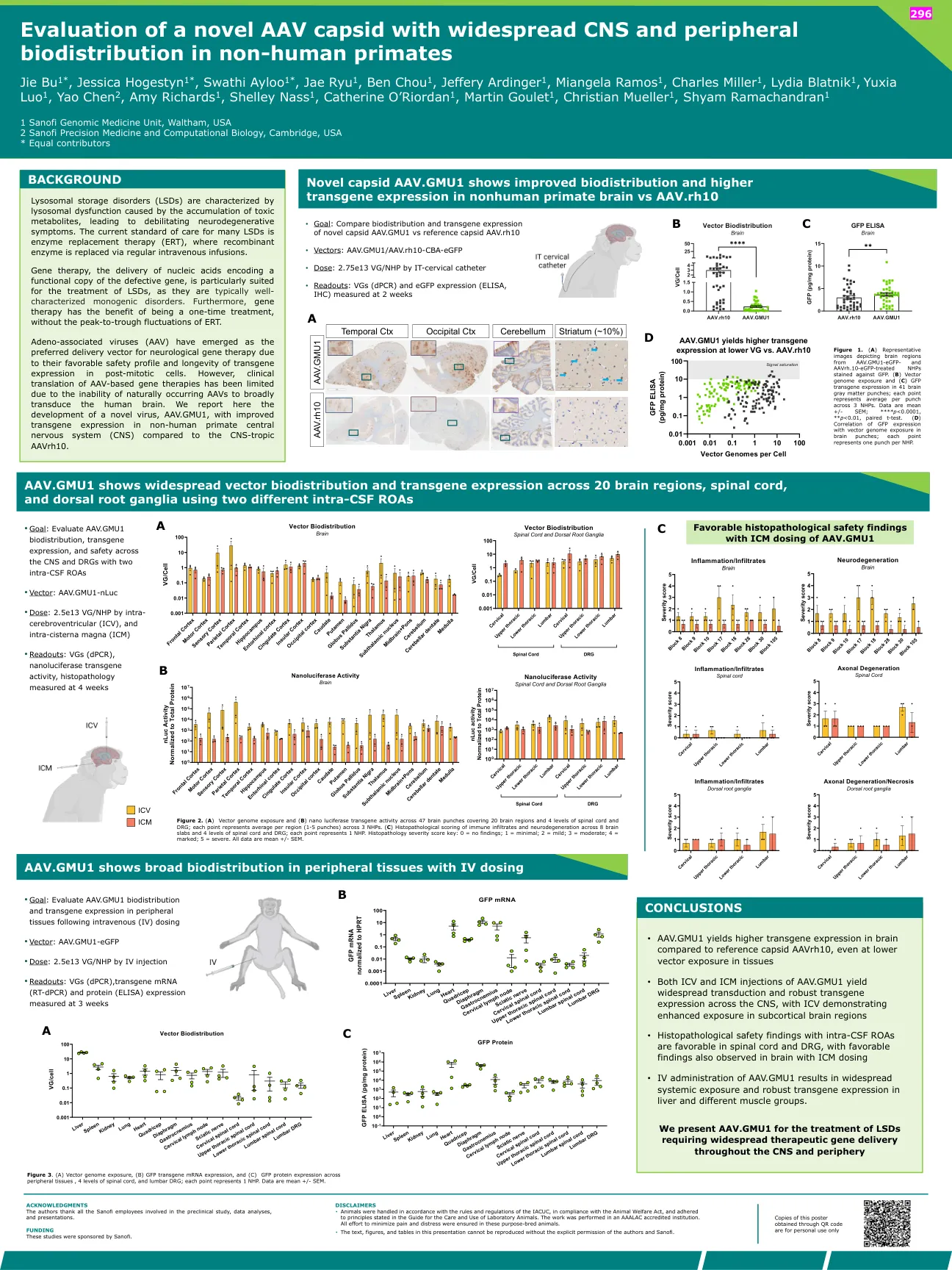
评估非人类灵长类动物的新型AAV CAPSID具有广泛的中枢神经系统和外围生物分布
与腺相关病毒(AAV)已成为神经基因治疗的首选递送载体,因为它们的安全性良好,并且在有丝分裂后细胞中转基因表达的寿命。然而,由于自然发生的AAV无法广泛传递人脑,因此基于AAV的基因疗法的临床翻译受到限制。我们在这里报告了一种新型病毒AAV.GMU1的开发,与CNS-Tropic AAVRH10相比,非人类灵长类动物中枢神经系统(CNS)的转基因表达改善。

解锁精度基因疗法:在CAPSID热点交换纳米机构
1。Wang,d。,Tai,P.W.L。 和gao,g。 (2019)腺相关病毒载体作为基因治疗递送的平台。 nat Rev Drug Discov 18,358-378。 2。 Jay,F.T。,Lughlin,C.A。 和Carter,B.J。 (1981)真核转化控制:腺相关的病毒蛋白合成受腺病毒DNA结合蛋白突变的影响。 Proc Natl Acad Sci U S A 78,2927-2931。 3。 Srivastava,A。,Lusby,E.W。 和Berns,K.I。 (1983)腺苷相关病毒2基因组的核苷酸序列和组织。 J Virol 45,555-564。 4。 Johnson,F.B。,Ozer,H.L。 和Hoggan,M.D。 (1971)腺病毒相关病毒的结构蛋白3. J Virol 8,860-863。 5。 Rose,J.A。,Maizel,J.V。,Inman,J.K。 和Shatkin,A.J。 (1971)腺病毒相关病毒的结构蛋白。 J Virol 8,766-770。 6。 Snijder,j。,van de Waterbeemd,m。,Damoc,e。,Denisov,e。,Grinfeld,d。,Bennett,A。,Agbandje-McKenna,M。,Makarov,A。 和Heck,A.J。 (2014)通过Orbitrap质谱定义了病毒和细菌纳米颗粒的化学计量和货物负荷。 J Am Chem Soc 136,7295-7299。 7。 xie,q。,bu,w。,bhatia,s。,hare,j。,somasundaram,t。,azzi,a。 和Chapman,M.S。 (2002)腺相关病毒(AAV-2)的原子结构,人类基因治疗的载体。 Proc Natl Acad Sci U S A 99,10405-10410。 8。 和Agbandje-Mckenna,m。Wang,d。,Tai,P.W.L。和gao,g。(2019)腺相关病毒载体作为基因治疗递送的平台。nat Rev Drug Discov 18,358-378。2。Jay,F.T。,Lughlin,C.A。 和Carter,B.J。 (1981)真核转化控制:腺相关的病毒蛋白合成受腺病毒DNA结合蛋白突变的影响。 Proc Natl Acad Sci U S A 78,2927-2931。 3。 Srivastava,A。,Lusby,E.W。 和Berns,K.I。 (1983)腺苷相关病毒2基因组的核苷酸序列和组织。 J Virol 45,555-564。 4。 Johnson,F.B。,Ozer,H.L。 和Hoggan,M.D。 (1971)腺病毒相关病毒的结构蛋白3. J Virol 8,860-863。 5。 Rose,J.A。,Maizel,J.V。,Inman,J.K。 和Shatkin,A.J。 (1971)腺病毒相关病毒的结构蛋白。 J Virol 8,766-770。 6。 Snijder,j。,van de Waterbeemd,m。,Damoc,e。,Denisov,e。,Grinfeld,d。,Bennett,A。,Agbandje-McKenna,M。,Makarov,A。 和Heck,A.J。 (2014)通过Orbitrap质谱定义了病毒和细菌纳米颗粒的化学计量和货物负荷。 J Am Chem Soc 136,7295-7299。 7。 xie,q。,bu,w。,bhatia,s。,hare,j。,somasundaram,t。,azzi,a。 和Chapman,M.S。 (2002)腺相关病毒(AAV-2)的原子结构,人类基因治疗的载体。 Proc Natl Acad Sci U S A 99,10405-10410。 8。 和Agbandje-Mckenna,m。Jay,F.T。,Lughlin,C.A。和Carter,B.J。(1981)真核转化控制:腺相关的病毒蛋白合成受腺病毒DNA结合蛋白突变的影响。Proc Natl Acad Sci U S A 78,2927-2931。3。Srivastava,A。,Lusby,E.W。和Berns,K.I。(1983)腺苷相关病毒2基因组的核苷酸序列和组织。J Virol 45,555-564。4。Johnson,F.B。,Ozer,H.L。 和Hoggan,M.D。 (1971)腺病毒相关病毒的结构蛋白3. J Virol 8,860-863。 5。 Rose,J.A。,Maizel,J.V。,Inman,J.K。 和Shatkin,A.J。 (1971)腺病毒相关病毒的结构蛋白。 J Virol 8,766-770。 6。 Snijder,j。,van de Waterbeemd,m。,Damoc,e。,Denisov,e。,Grinfeld,d。,Bennett,A。,Agbandje-McKenna,M。,Makarov,A。 和Heck,A.J。 (2014)通过Orbitrap质谱定义了病毒和细菌纳米颗粒的化学计量和货物负荷。 J Am Chem Soc 136,7295-7299。 7。 xie,q。,bu,w。,bhatia,s。,hare,j。,somasundaram,t。,azzi,a。 和Chapman,M.S。 (2002)腺相关病毒(AAV-2)的原子结构,人类基因治疗的载体。 Proc Natl Acad Sci U S A 99,10405-10410。 8。 和Agbandje-Mckenna,m。Johnson,F.B。,Ozer,H.L。和Hoggan,M.D。(1971)腺病毒相关病毒的结构蛋白3.J Virol 8,860-863。5。Rose,J.A。,Maizel,J.V。,Inman,J.K。 和Shatkin,A.J。 (1971)腺病毒相关病毒的结构蛋白。 J Virol 8,766-770。 6。 Snijder,j。,van de Waterbeemd,m。,Damoc,e。,Denisov,e。,Grinfeld,d。,Bennett,A。,Agbandje-McKenna,M。,Makarov,A。 和Heck,A.J。 (2014)通过Orbitrap质谱定义了病毒和细菌纳米颗粒的化学计量和货物负荷。 J Am Chem Soc 136,7295-7299。 7。 xie,q。,bu,w。,bhatia,s。,hare,j。,somasundaram,t。,azzi,a。 和Chapman,M.S。 (2002)腺相关病毒(AAV-2)的原子结构,人类基因治疗的载体。 Proc Natl Acad Sci U S A 99,10405-10410。 8。 和Agbandje-Mckenna,m。Rose,J.A。,Maizel,J.V。,Inman,J.K。和Shatkin,A.J。(1971)腺病毒相关病毒的结构蛋白。J Virol 8,766-770。6。Snijder,j。,van de Waterbeemd,m。,Damoc,e。,Denisov,e。,Grinfeld,d。,Bennett,A。,Agbandje-McKenna,M。,Makarov,A。和Heck,A.J。(2014)通过Orbitrap质谱定义了病毒和细菌纳米颗粒的化学计量和货物负荷。J Am Chem Soc 136,7295-7299。7。xie,q。,bu,w。,bhatia,s。,hare,j。,somasundaram,t。,azzi,a。和Chapman,M.S。(2002)腺相关病毒(AAV-2)的原子结构,人类基因治疗的载体。Proc Natl Acad Sci U S A 99,10405-10410。8。和Agbandje-Mckenna,m。Govindasamy,L。,Padron,e。,McKenna,R.,Muzyczka,n。,Kaludov,n。,Chiorini,J.A。(2006)在结构上绘制腺相关病毒血清型4的多种表型。J Virol 80,11556-11570。9。tse,l.v。,Klinc,K.A。,Madigan,V.J。,Castellanos Rivera,R.M。,Wells,L.F。,Havlik,L.P。,Smith,J.K。和Asokan,a。(2017)结构引导的抗原不同的腺相关病毒变体用于免疫逃避。Proc Natl Acad Sci U S 114,E4812-E4821。10。Chan,K.Y。,Jang,M.J。,Yoo,B.B.,Greenbaum,A。Chan,K.Y。,Jang,M.J。,Yoo,B.B.,Greenbaum,A。

无细胞试验表明 HIV-1 衣壳可保护逆转录产物免受 cGAS 免疫感应的影响
逆转录病毒可被先天免疫传感器环鸟苷酸环磷酸腺苷合酶 (cGAS) 检测到,该合酶可识别逆转录 DNA 并激活抗病毒反应。然而,HIV-1 保护其基因组免受 cGAS 识别的程度仍不清楚。为了详细研究这一过程的机制,我们在无细胞系统中重建了 HIV-1 的逆转录、基因组释放和先天免疫感应。我们发现,即使在完成逆转录后,野生型 HIV-1 衣壳也能保护病毒基因组免受 cGAS 的侵害。病毒 DNA 可能因热应激、衣壳突变或肌醇六磷酸 (IP6) 浓度降低而“脱保护”,这些因素会使衣壳不稳定。令人惊讶的是,衣壳抑制剂 lenacapavir 也会破坏病毒核心并显著增强 cGAS 活性,无论是在体外还是在细胞感染中。我们的研究结果提供了生化证据,表明 HIV-1 衣壳晶格隐藏了 cGAS 的基因组,而病毒核心的化学或物理破坏可以暴露 HIV-1 DNA 并激活先天免疫信号。

AAV CAPSID的可变区域I中的单个氨基酸变体赋予肝脏脱落
。cc-by-nc-nd 4.0国际许可证(未经同行评审证明)获得的是作者/资助者,他授予Biorxiv授予Biorxiv的许可,以永久显示预印本。这是该版本的版权持有人,该版本发布于2025年3月5日。 https://doi.org/10.1101/2025.03.04.641478 doi:Biorxiv Preprint

开发AAV-GBA1基因替代疗法,用于通过血液屏障渗透AAV CAPSID
a-b)VG生物分布是通过DDPCR,FAM-RBGPA和VIC-MSTFRC探针来测量二倍体动物细胞的。在皮质(CTX)和丘脑(Th)中证明了跨前脑和中脑区域的成功基因转移。最小基因转移(<1VG/细胞)。d -f)GBA1 mRNA表达。用于该测定法的7个PLEX探针集是定制的,旨在区分转基因特异性GBA mRNA和小鼠内源性GBA mRNA。mRNA表达值据报道为平均荧光强度(MFI)。人类GBA1 mRNA如图d -f所示。在皮质和丘脑中证明了跨大脑区域的成功转录。在肝脏中观察到降低的表达。g - i)使用Sensolote®蓝葡萄糖脑培合酶活性测定法测量皮质,丘脑和肝脏的Gcase活性。高剂量的Php.eb.gba1具有增强10、2、5和3的高剂量,在中枢神经系统组织中的GCASE活性显着增加。平均值+/- SEM。
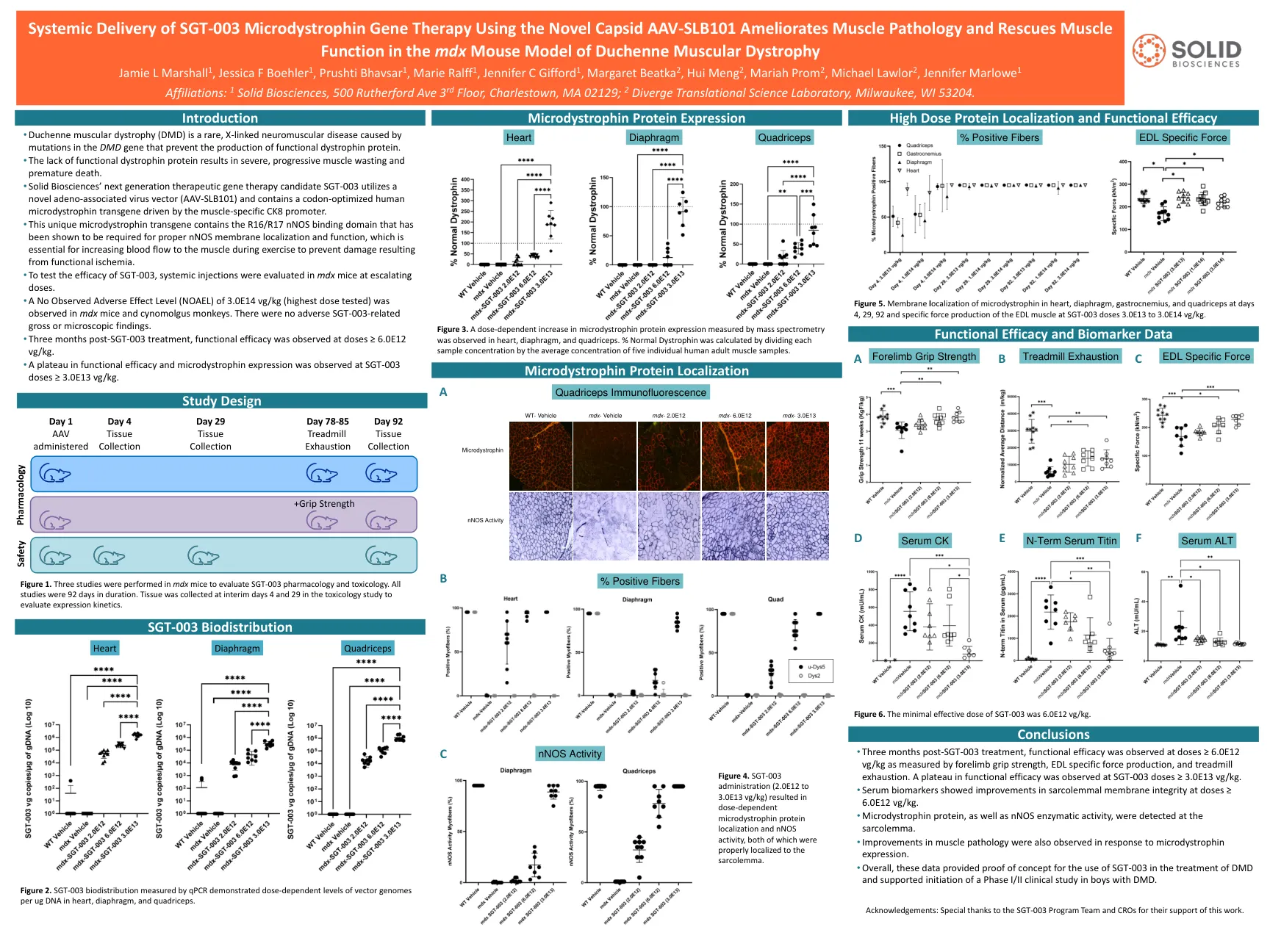
使用新型的CAPSID AAV-SLB101的SGT-003微型肌营养物基因疗法的全身递送可改善肌肉病理学并挽救肌肉
被证明是适当的NNOS膜定位和功能所必需的,这对于在运动过程中增加血液流向肌肉以防止功能性缺血引起的损害至关重要。•为了测试SGT-003的功效,在MDX小鼠中评估了全身注射

表征与液相色谱质谱法(LC-MS)的肽相关病毒(AAV)衣壳蛋白的基于液相色谱(LC-MS)的肽映射和翻译后修饰分析(PTMS)
肽映射样品制备:AAV8参考材料在2x10 13 Vg/ml的浓度下包含20μl的总体积。这导致消化的估计总蛋白浓度为0.12μg/μL,总蛋白质为2.4μg。将AAV样品在6 m尿素中变性,在80℃以1 mm DTT变性30分钟,然后用15 mm iodoacetamide烷基化在黑暗中的室温下在室温下30分钟。将还原和烷基化的样品冷却至室温,并用3次同等体积的缓冲液(50 mM Tris-HCl和1 mm CaCl 2 [pH 7.5])稀释,将尿素浓度降低至<2M。然后将样品降低到<2M。然后用0.4 µ µGGGRYPSIN或CHYMOTRYPESIN或CHYMOTRYPSIN或CHYMOTRYPRYRYPERSIN或CHYMOTRYPRYRYPRYRYPRYRYPRYRYSIL逐夜消化。通过将甲酸添加到最终浓度的10%中终止消化,并将样品直接注入LCMS-9050进行分析。

Martin Rhee MD,吉利德(Gilead
Martin Rhee MD,Gilead“ Gilead La/Er Pipeline的当前地位” Len(小分子Capsid抑制剂)的执行主任是Gilead以人为本的La Portfolio的基础。有利的特征。•高效力(EC50 = 100pm);低剂量需求,并具有延长的给药间隔。•多模式机制;现有化合物没有重叠电阻的第一类代理。•特征良好的临床PK,包括长半衰期。•柔性给药pro(口服或注射剂量)。len作为HIV治疗的组合疗法 - 跨药物和给药途径的各种选择。管道。•注射式(预先为p3):Q3M给药是直接的重点; Q6M剂量是长期目标。o Q6M化合物(Pre-Ind):GS-1219(Insti)和GS-3242(Insti)。o Q3M化合物(P1):GS-6212(Insti)和GS-1614(NRTTI; Merck-Gilead Collaboration)。o Q6M完整方案(P2):Len + 2bnabs(Terapavimab [Tab]和Zinlirvimab [Zab])。•口服(P1至P2):QW剂量是直接的重点。o QW化合物(P1):GS-1720(Insti)和GS-4182(Len Prodrug)。o QW完整方案(P2):LEN + ISL(NRTTI; Merck-Gilead Collaboration)。o QD完整方案(P2):LEN + BIC(Insti)。完整方案的临床发展。•Q6M注射Len SC + 2 BNABS IV(TAB + ZAB)。o P1B研究(LEN + 2bnabs [10mg/kg和30mg/kg])在VS PWH中具有病毒易感性对Tab和Zab的敏感性(n = 21)。*两个剂量组在第26周保持病毒学抑制(Eron J等人HIV 2024)。o小型P1B Pilot队列研究与PWH相对于PWH,病毒易感性仅对一个BNAB(n = 10)。*结果将在CROI 2024(口头#2258; Eron J等)上呈现。o正在进行的P2随机(2:1)研究(LEN + 2bnabs vs基线ART)与PWH中具有病毒易感性对TAB和ZAB的敏感性(n = 75);在第26和52周的安全性和效率终点。*完全注册。•QW口服ISL + Len(Merck-Gilead Collaboration)。o P2随机(1:1)研究(ISL + LEN与QD B/F/TAF)在B/F/TAF上的PWH中(n = 100)。*由于ISL淋巴细胞影响,2021年FDA临床保留;研究在2023年恢复使用下ISL剂量(2mg)。*第24周的安全性和效率将在CROI 2024(Late-Breaker#208; Colson A等人)。•QD口服BIC + LEN(不是LA;旨在满足复杂方案上的VS PWH的需求)。o正在进行的P2随机(2:2:1)开关研究(QD口服BIC + LEN [25mg或50 mg] vs基线多平板电脑方案)。*结果将在CROI 2024(海报#1289; Mounzer K等人)中呈现。o刚开始P3随机(2:1)研究(BIC [75mg] + LEN [50 mg] vs基线多磁盘方案)。len作为预防艾滋病毒的Q6M单一治疗 - 潜力解决日常药丸的负担/污名并增加了全球PREP的吸收。目的程序。•五项研究故意旨在招募临床和预防研究的人口:妇女,尤其是青少年女孩和年轻妇女(AGYW);变性人(TG);和性别非二进制(GNB)。•概念证明:衣壳抑制剂可防止NHP中的猿猴艾滋病毒;有和没有艾滋病毒的人的强大安全性和PK数据库。•杠杆:伙伴关系;可能受益于准备和社区的人的意见;以人为本的设计;以及多样性公平与包容(DEI)。

Ravula Tulasi Naga Pavan Kumar,Int。 J. of Pharm。 Sci。,2024,第2卷,第11期,961-971
人类免疫缺陷病毒1型(HIV-1)仍然是全球健康挑战,尽管抗逆转录病毒疗法的进步。衣壳抑制剂已成为有前途的药物,因为它们的独特作用机理靶向病毒capsid,该机制在病毒复制,组装和不涂层中起着关键作用。在其中,Lenacapavir(一种第一类长效的衣壳抑制剂)在临床前和临床试验中显示出很大的疗效,表明抗病毒活性延长,给药频率降低,并且安全性良好。本评论探讨了新型衣壳抑制剂的药理学,临床功效和安全性,重点是Lenacapavir解决依从性和耐药性问题的潜力。此外,本文讨论了CAPSID抑制剂研究中的实际考虑,给药策略以及未来的方向,为下一代HIV疗法提供了见解。