XiaoMi-AI文件搜索系统
World File Search Systemprogression

CMF 35 进度表
GEOINT 图像分析师利用、解释、分析和传播来自国家、战区、无人值守地面传感器和战术系统的图像,为作战提供 IMINT 支持,并且图像分析师可以分析图像以识别和传递环境危害、民事考虑和威胁活动到在战场上机动的部队;通过基线或中间图形、战斗损伤评估和附带损伤估计为致命和非致命目标提供支持。

CMF 42A 进阶图表
最合格的资格如下: SSG :成功担任营(或同等级别)S1 SR HR SGT 18-24 个月,并且在扩大任务中成功担任 12 个月,具有 MQ 评级和高度计算排名 SFC :成功担任营/旅 S1 SR HR SGT 18-24 个月,目前或之前为 NCO 等级(评估表明为 SSG 或以上),并且在扩大任务中成功担任 12 个月,具有 MQ 评级和高度计算排名 MSG/1SG:成功担任旅 S1 SR HR SGT 18-24 个月,目前等级,并且成功担任 1SG 12 个月,或在扩大任务中具有 MQ 评级和高度计算排名 CSM/SGM :担任师 G1 SGM 18-24 个月;陆军服务组件司令部 G-1;军团级 G1 士官长;具有 MQ 等级和高数字排名的营/旅 CSM

CMF 92 进度表
最符合条件的如下: SSG:优异的学业成绩;杰出的荣誉毕业生;指挥官名单;副学士学位;高级士兵联合专业军事教育 I(SEJPME I);认证石油专家 3 级 SFC:优异的学业成绩;杰出的荣誉毕业生;指挥官名单;学士学位;高级士兵联合专业军事教育 II(SEJPME II);认证石油专业人员 5 级 MSG:优异的学业成绩;杰出的荣誉毕业生;指挥官名单;硕士学位;高级士兵联合专业军事教育 II(SEJPME II);认证石油专业人员 5 级 CSM/SGM:优异的学业成绩;硕士学位
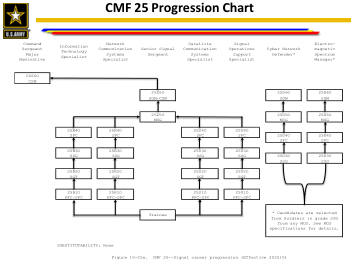
CMF 25 进度表
最合格的看起来像: SSG:杰出荣誉毕业生/荣誉毕业生;30 多个大学学分;信息保证技术员 II 级/信息保证管理 I 级认证、CompTIA 网络安全分析师、EC-Council 认证网络防御者、认证道德黑客、微软认证解决方案专家 SFC:杰出荣誉毕业生/荣誉毕业生;副学士学位;信息保证技术员 III 级/信息保证管理 II 级认证、认证道德黑客、思科认证网络专业人员、GIAC 认证取证分析师、微软认证解决方案专家 MSG:杰出荣誉毕业生/荣誉毕业生;学士学位;信息保证技术员 III 级/信息保证管理 III 级认证、认证道德黑客、认证信息系统审计师、GIAC 系统和网络审计师、微软认证解决方案专家 SGM:硕士学位;信息保证管理 III 级、认证信息系统安全专业人员、认证信息安全经理、认证首席信息安全官、微软认证解决方案专家

CMF 36 进度表
培训 最合格的培训课程如下: SSG:教官学校;招聘人员;战斗顾问培训课程;空降;空中突击;士兵助手培训课程;士兵联合专业军事教育 (EJPME) I;平等机会领导者;通用教师发展 - 讲师课程;通用教师发展 - 开发者课程;战斗参谋 SFC:监察长课程;通用教师发展 - 讲师课程;EJPME II;战斗参谋;陆军主计长课程;执行主计长课程;通用教师发展 - 开发者课程 MSG:通用教师发展 - 讲师课程;中级数据分析课程 CSM/SGM:营/旅指挥前课程;执行主计长课程;高级商业分析认证;高级资源经理课程

非攻击性冠状动脉斑块的进展
gianluca pontone 1,2·Alexia Rossi 3,4·Andrea Baggiano 1·Daniele Andreini 1,5·Edoard 7·Marco Guglielmo 1·Giuseppe Muscogiuri Muscogiuri 1·saima Mushtaq 1·saima Mushtaq 1·MotOult J.MOUGAH S. 10·Filippo Cademartiri 11·Kavitha Chinnaiyan 12 Jung Hyun Choi 13·Eun Ju Chun 14·Pedro dearújoGonçalves15,16·ilan Gottlieb 17·Martin Hadamitzky 18·Yong Jin Kim 19·Yong Jin Kim 19·Yong jin Kim byoun byun lee lee lee 20·lee 21 21 23 · Hugo Marques 15 · Habib Samady 24 · Sanghoon Shin 21 · Ji Min Sung 22,25 · Alexander Van Rosendael 26 · Renu Virmani 27 · Jeroen J. Bax 28,29 · Jonathon A. Leipse 30 · Fay Y. Lin 26 · James K. Min 31 · Jagat Narula 32 · Leslee J. Shaw 26 · Hyuk -Jae Chan 22,25

DNA键入标记的进展
•五个RFLP探针提供了几乎独特的身份(〜1 in 10 9个个体)•RFLP需要至少25 ng相对未依赖的DNA(1000-20,000底发)•短串联重复序列(strs)仅需要〜1 ng DNA,只需〜1 ng dna即可降级•辨别力•5 rflp probient•rflp probient equi fim

学徒制进阶路线列表
农业和园艺 1 树艺 L 6 2 年 戈尔韦和罗斯康芒 ETB - 晋升路线 - 树艺职业道路 - 学徒 - 合格树艺师 - a) 继续教育晋升(树艺或园艺资格) - 专业培训师/教师 - b) 地方当局市/县议会(团队负责人)- 树艺师/树木官员 - c) 商业私营公司(团队负责人) - 公司所有者/顾问。 2 农场经理 L 7 2 年 Teagasc - Teagasc 建议他们将在未来几个月探索晋升选项,毕业生可能会提前进入 8 级课程。 3 农场技术员 L 6 2 年 Teagasc - 晋升至 - 农场经理学徒(专业可持续农场管理普通学士学位 - NFQ 7 级)。 4 园艺 L 6 2 年 Teagasc - 高级进入东南技术大学 7 级普通园艺学士学位。

Switch 进程路线图
对于许多有特殊需求的儿童,AT 始于多感官室,其中使用泡泡管、轮式投影仪、镜球和音乐等设备提供听觉和视觉刺激,通常还伴有手部和足部按摩,以帮助儿童发展本体感受意识。一些学校在多感官室安装了数字投影仪或交互式等离子屏幕,使儿童能够使用电脑和软件,将多感官体验扩展到包括照片、动画和视频。

2024-2025学生进步计划
背景和目的路易斯安那州法律(R.S.17:24.4)要求当地教育机构(LEAS)根据路易斯安那州教育评估计划(LEAP)的学生绩效建立全面的学生进步计划(PPP),并具有与路易斯安那州能力教育计划兼容的目标和目标,并根据基于路易斯安那州的教育计划兼容,并根据基于基本委员会的最低限度委员会和次要教育(BESE)的最低标准(BESE)。该计划应解决学生的安置和晋升,并需要学生掌握适合等级的技能,然后才能推荐他或她进行晋升。法律规定:“应特别强调学生在晋升和安置中可以考虑的成绩技能的熟练程度;但是,每个地方学校董事会均应制定有关学生专业运动和安置的政策。”该法律继续要求路易斯安那州教育部(LDOE)建立未经BESE的批准,即在数学,英语艺术,科学和社会研究中的四年级和八年级LEAP考试中的成就水平,学生需要使学生晋升为五年级和九年级。BESE还必须确定当学生无法符合董事会批准的最低学术标准时,要使用各种干预选项的性质和应用。本文件的目的是协助LEA根据适用的法律和法规开发其所需的PPP,并将与学生安置和晋升有关的LEA政策和程序编纂。有关此文档的问题应引向ppp@la.gov在本文档的每个部分中,符合适用法律和法规的语言已经预先填充。空间,以添加任何其他本地政策和程序,以履行法律的授权,并支持学生获得熟练技能的熟练程度。一旦完成,提交给LDOE并在当地出版,教师应在个人基础上确定每个学生的晋升或安置。leas可以审查晋升和安置决策,以确保遵守其既定政策,并且可以由学校的理事机构,当地校长或学生的父母或法律保管人发起审查。