XiaoMi-AI文件搜索系统
World File Search Systemprokaryotes


维生素B12并非所有海洋原始细菌与其环境共享
维生素B 12(钴胺,本文B 12)是大多数原核,真核微生物和动物的氨基酸合成和碳补给TCA循环的必不可少的辅助因。尽管大多数人都要求,但B 12仅由少量的原核生物产生,因此导致原子营养和辅助营养物之间的复杂相互作用。然而,未知B 12是如何由概念营养提供的。在这项研究中,在与Thalassiosira pseudonana(A B 12富营养硅藻)的共同培养中生长了33 B 12个原型型植物菌株,以确定通过共享B 12来支持硅藻生长的细菌能力。被鉴定为与硅藻共享B 12,而9个被鉴定为保留B 12,并且不支持硅藻的生长。其他细菌仅在添加底物的情况下与硅藻共享B 12,或者抑制了硅藻的生长。细胞外B 12测量B 12 -provider和b 12-取消菌株证实,只能在经过测试的B 12 -provider菌株的环境中检测到辅助因子。通过LC -MS测量细胞内B 12,并表明不同B 12-构造剂和B 12的retainer菌株的浓度有很大不同。尽管B 12对于绝大多数微生物至关重要,但导出该基本辅助因子的机制仍然未知。我们的结果表明,可以合成b 12 de Novo的大部分细菌不能与环境共享辅助因子。

外源 DNA 序列的“同类相食”:适应性免疫的祖先形式,需要识别危险
适应性免疫是一种复杂的免疫反应形式,能够保留大量靶抗原(表位)作为非自身抗原的分子记忆。当它再次遇到具有已知表位的免疫球蛋白或 T 细胞受体抗原结合位点时,它能够重新激活自身,而这些表位之前曾激活过宿主免疫系统。长期以来,人们一直认为适应性免疫是一种高度进化的非自身识别形式,在物种形成过程中出现得相当晚,是对一种更普遍的反应(称为先天免疫)的补充。先天免疫提供了一种相对非特异性的防御(尽管由能够特异性识别病毒或细菌化合物的传感器介导),并且不保留对危险的记忆。但是,这种最近获得适应性免疫的概念受到了挑战,因为原核生物中已经存在另一种形式的特定识别机制,这种机制可能能够特异性地自动防御外部危险。这种识别机制可以被认为是一种原始形式的特定(适应性)非自身识别。它基于这样一个事实:许多古细菌和细菌使用一种基因组编辑系统,该系统赋予原核生物适当的病毒 DNA 序列的能力,使它们能够通过一种与适应性免疫非常相似的机制来防止宿主受到损害。这被模糊地称为“外来 DNA 的内源化”或“病毒 DNA 捕食”,或者更形象地说是“DNA 同类相食”。多年来,证据不断积累,突显了外来 DNA 的内源化在与适应性免疫相关的基本过程中的关键作用,并导致了适应性免疫在物种形成后期出现的教条的改变。

剪接背景:转录是使用...
DNA一次从3'末端到5'末端读取一个底座。对于每个读取的碱基,核苷酸都配对以生长RNA链。这一直持续到转录终止为止。在原核生物中,该新的RNA转录本可用于翻译。在真核生物中,这种新制成的RNA链称为前MRNA转录本。在可以用于翻译之前,必须进行转录后修改以使其成为成熟的mRNA转录本。在阅读之前,停止并回答以下思想问题以测试您的当前知识:前MRNA转录本与成熟的mRNA转录本有何不同(要具体)?

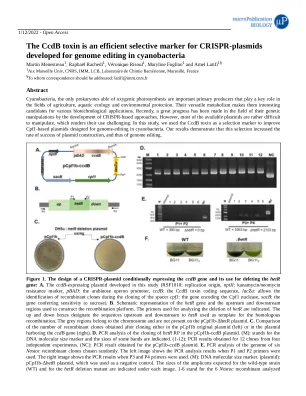
CcdB 毒素是 CRISPR 的有效选择标记...
蓝藻是唯一能够进行产氧光合作用的原核生物,是重要的初级生产者,在农业、水生生态和环境保护领域发挥着关键作用。它们多功能的代谢使它们成为各种生物技术应用的有趣候选者。最近,通过基于 CRISPR 的方法的发展,它们的基因操作领域取得了巨大进展。然而,大多数可用的质粒都很难操作,这使得它们的使用具有挑战性。在本研究中,我们使用 CcdB 毒素作为选择标记来改进用于蓝藻基因组编辑的基于 Cpf1 的质粒。我们的结果表明,这种选择提高了质粒构建的成功率,从而提高了基因组编辑的成功率。

BME-生物医学工程
BME 464/564 低温等离子体的生物医学应用(3 学分)本课程与 ECE 和生物学交叉列出。它旨在供高年级本科生和一年级研究生选修。课程内容是多学科的,结合了工程学和生物科学的材料。本课程涵盖非平衡等离子体、低温等离子体源和细胞生物学的基础知识。随后详细讨论低温等离子体与生物细胞(原核生物和真核生物)的相互作用。将涵盖医学中的潜在应用,例如伤口愈合、血液凝固、灭菌和杀死各种类型的癌细胞。先决条件:高年级
CcdB 毒素是 CRISPR 的有效选择标记...
蓝藻是唯一能够进行产氧光合作用的原核生物,是重要的初级生产者,在农业、水生生态和环境保护领域发挥着关键作用。它们多功能的代谢使它们成为各种生物技术应用的有趣候选者。最近,通过基于 CRISPR 的方法的发展,它们的基因操作领域取得了巨大进展。然而,大多数可用的质粒都很难操作,这使得它们的使用具有挑战性。在本研究中,我们使用 CcdB 毒素作为选择标记来改进用于蓝藻基因组编辑的基于 Cpf1 的质粒。我们的结果表明,这种选择提高了质粒构建的成功率,从而提高了基因组编辑的成功率。

PRPE,一种来自枯草芽孢杆菌的PPP蛋白磷酸酶,具有异常的底物
蛋白质磷酸化或去磷酸化是在所有生物体中发现的信号传递的重要机制。多年来,蛋白激酶和磷酸酶的性质被认为在原核生物和真核生物中是不同的。证明主要发生在组氨酸和天冬氨酸残基上,而相反,通常在丝氨酸,苏氨酸或酪氨酸残基上修饰真核蛋白。然而,近年来在细菌中报道了真核样蛋白激酶和磷酸酶,相反,在真核生物中发现了原核性蛋白质的ASP酶的同源物(有关评论,请参见[1-7])。这些研究表明,真核生物和原核生物可能具有所有类型的信号转导的相似机制。蛋白磷酸酶可以根据其酶特异性(即促磷酸酶和Tyr磷酸酶)分为两组[8,9]。ser} THR磷酸酶在ITRO中显示出广泛的特异性,并已分为四类:PP1,PP2A,PP2B和PP2C,根据保守的基序,它们对抑制剂和离子的抑制剂和离子需求的敏感性[9-11]。氨基酸序列比较表明PP1,PP2A和PP2B是同一PPP家族的成员[10]。PPP家族代表了较高的真核生物中蛋白质ser}的最大蛋白质ser} [12]。这些酶还与对称的折断氨酸四磷酸酶具有序列相似性[13]。被识别的PPP家族的第一个原核生物是噬菌体λ221的乘积[14]。目前,几个成员在ARCHEA和细菌中均已详细介绍[15-19]。但是,关于生理学的数据很少
分子生物学(3cfu)综合课程
超螺旋和拓扑性质。拓扑异构酶。细菌类核。组蛋白和核小体的性质和组装。染色质的高级结构。组蛋白的翻译后修饰。溴多胺和染色质结构域。表观遗传学。原核生物和真核生物的基因组。复制模型。DNA合成。细菌DNA聚合酶。校对和缺口翻译。复制子模型。OriC和半甲基化。Ter/Tus。真核细胞核中的复制工厂。ARS结构和复制控制。酶学。前RC和前启动复合物。复制抑制剂,如化疗药物和抗病毒药物。端粒和端粒酶的结构、功能和意义。DNA损伤和修复。基因组作为动态实体。体细胞和种系突变。SNP。内在和外在损伤。化学和物理诱变剂。原核生物和真核生物中的去除、逆转和损伤避免系统。MUT 系统。BER 系统。糖基化酶的重要性。安全系统。NER 系统:UvrABCD 和 XP 蛋白。GG-NER 和 TC-NER。光解作用、MGMT、AlkBH。损伤耐受机制。TLS。细菌中的 SOS 反应。单丝和双丝断裂。HR 和 NHEJ。由于修复系统突变而导致的人类疾病。位点特异性重组。重组酶。Lambda 噬菌体。Cre-Lox 系统和 KO 小鼠。简单和复杂的转座子。SINE 和 LINE 元素、Alu 序列。原核生物和真核生物中的 RNA。结构、类型和特性。细菌 RNA 聚合酶和相关因子。转录单位。转录步骤。细菌启动子中的共识序列。终止机制。抑制剂。 Lac、ara 和 trp 操纵子。阳性和阴性对照。真核细胞中的 RNA 类别。RNA 聚合酶 (CTD) 的结构和功能。三种启动子的特征。基础转录机制。TFIIH。反式激活因子、辅激活因子。CpG 岛甲基化。组蛋白密码。长程调节剂。DNA 结合蛋白的功能域 (HTH、HD、HLH、ZF、LZ)。RNA 成熟、核运输和转录后控制。加帽类型。添加 polyA。CTD 的变化。外显子和内含子。外显子改组。四类内含子及其去除机制。剪接体和剪接位点。AT-AC 剪接。EJC 复合体。可变剪接。ESE 和 ESS 序列、SR 和 hnRNP 蛋白。SMN 基因。剪接和病理。rRNA 和 tRNA 加工反应。核糖体基因。 SnoRNA 和核仁功能。RNA 编辑。插入和转换编辑。人类 RNA 编辑的示例。细胞核和细胞质中的 RNA 周转。外泌体。无义介导的 mRNA 衰变 (NMD)。非编码 RNA。小 RNA 在细胞中的功能。RNA 干扰。siRNA。微小 RNA 的生物发生。miRNA、长链非编码 RNA、环状 RNA 的作用机制。逆转录病毒的一般信息。遗传密码和翻译。遗传密码的性质和特征。线粒体密码。ORF。tRNA 的特征。不常见碱基。aa-tRNA 合成酶的功能和类别。遗传密码的翻译重编码和扩展。SeCys。核糖体是一种核酶。原核生物和真核生物的翻译阶段。不同的启动机制。能量成本。NSMD。细菌中的 tmRNA。抑制剂。蛋白质的翻译后修饰、分选和降解。折叠和错误折叠。朊病毒。HSP60 和 HSP70。泛素和泛素化系统。SUMO 化糖基化。蛋白酶体。肽信号。蛋白质分选。线粒体输入。线粒体基因组细胞中的线粒体可塑性。人类线粒体基因组。遗传、结构、复制及其表达的原理。线粒体 DNA 中的改变。DNA 克隆的原理。修饰限制系统。克隆载体。cDNA 合成。基因组 DNA 和 cDNA 文库。TA 克隆。表达克隆。基因表达沉默。基因治疗。数据库。基因组编辑元件(Talen、Zn 指、CRISPR/Cas9 系统)。PCR 和 DNA 测序。PCR 的特性。PCR-RFLP。实时 PCR、DNA 测序。NGS。核酸杂交。杂交原理。熔点和严格性。探针制备:切口平移。Southern、Northern、杂交测定。蛋白质印迹。
