XiaoMi-AI文件搜索系统
World File Search System冯永真

ecotin:一种多功能细菌和真核生物的蛋白酶抑制剂
丝氨酸蛋白酶抑制剂是参与重要途径和过程的大型蛋白质,例如炎症反应和血液凝结。大多数以精确的作用方式为特征,从而靶向狭窄的蛋白酶底物。然而,丝氨酸 - 聚糖抑制剂Ecotin能够抑制广泛的丝氨酸蛋白酶,这些丝氨酸蛋白酶显示出广泛的特异性。这种特异性是由特殊的结构特征驱动的,这些特征允许在与目标结合时独特的灵活性。尽管在许多人/动物相关的细菌中经常观察到,但在植物相关的分类单元和环境物种中也可以发现Ecotin同源物。本综述的目的是提供有关生物学重要性,在宿主 - 微生物相互作用中的作用以及从整个生命树中的真核和原核物种分离的生态蛋白直系同源物之间的进化关系。

真核生物FES解放酶的综合结构方法
silvia.onesti@elettra.eu解旋酶是必不可少的,无处不在的酶,在各种细胞过程中起着关键作用,从DNA复制到修复,重组以及RNA翻译和运输。由于它们在病毒,细菌和真核细胞中的重要作用,它们正成为一类新的抗菌,抗病毒和蚂蚁癌药物靶标。通过解决/重塑各种非典型的DNA结构(例如G-四链体,Triplexe,holliday连接器,以及流离失所环(D-ROOPS和R-Loops))来发挥专业和特定功能:在这些主要作用中,有两个家族由Helicases of Helicases of Helicases of Helicases of Helicases formals of Family,扮演的是helicase of Helicases famessemass famesse formals formemase forme of Helicase,Floop femers of Helicases,Floops。含有FES群体的解旋酶无处不在,但其确切的作用机理知之甚少。特别是,对于FANCJ,DDX11和RTEL1,没有任何与医学上的与医学上的成员相关的结构信息。固有构象柔韧性,FES群集的稳定性和大小的结合使它们具有挑战性的结构生物学目标。

基于CRISPR的测定法,用于研究真核DNA维修的国际空间站
在我们探索地球以外的探索时,宇航员可能会面临因电离辐射引起的有害DNA损害的风险。双链断裂是一种可以通过两种主要的细胞途径来修复的DNA损伤:非同源末端连接,在此期间可以在断裂部位添加插入或插入,并同源重组,其中DNA序列通常保持不变。先前的工作表明,空间条件可能会影响DNA修复途径的选择,从而有可能使太空旅行期间辐射增加的风险增加。但是,我们对这个问题的理解受到技术和安全问题的限制,这些问题阻止了对太空中DNA修复过程的整体研究。CRISPR/CAS9基因编辑系统为真核生物中的双链破裂提供了一个模型。在这里,我们描述了一种基于CRISPR的基于CRISPR的测定法,用于完全在空间中选择双链破裂修复途径的评估。在此过程中必要的步骤中,我们描述了空间中第一个成功的遗传转化和CRISPR/CAS9基因组编辑。这些里程碑代表了国际空间站的分子生物学工具包的显着扩展。


利用热噪声理论和真随机数生成实现后量子密码学
量子计算机和算法的出现对对称和非对称密码系统的语义安全性提出了挑战。因此,实现新的密码原语至关重要。它们必须遵循量子计算器的突破和特性,因为量子计算器使现有的密码系统变得脆弱。在本文中,我们提出了一个随机数生成模型,该模型基于对体积为 58.83 cm 3 的电子系统体积元素热噪声功率的评估。我们通过对每个体积元素的温度进行采样来证明攻击者很难进行攻击。在 12 秒内,我们为 7 个体积元素生成一串 187 位随机密钥,这些密钥将通过量子密码学的特性从源传输到目的地。

具有双果冻的真核DNA病毒的自然史 -
门前病毒(Kingdom Bamfordvirae,Realm varidnaviria)是多种病毒的广泛组合,其相对较短的双链DNA基因组(<50 kbp)产生了由双果冻 - 双果冻 - 卷胶卷蛋白构建的二十os虫。前肿瘤动物感染所有细胞结构域的宿主,证明其古老的起源,尤其是与真核生物的七个超级组中的六个有关。前肿瘤分子包括四个主要的病毒组,即Polinton,Polinton,例如病毒(PLV),病毒噬细胞和腺毒。我们使用蛋白质结构建模和分析来表明蛋白质的DNA聚合酶(PPOLBS),polins,病毒噬细胞和细胞质线性质粒涵盖了n-终末结构域与末端蛋白(TPS)的N-末端域同源物(TPS),例如原始prd1-涉及tpectiricotic andototic artectirIdotics和eukaryotic artirIdotics artirIdotic artirIdotic artineciridotics anden tectirifiridotic toNERIFIRIDICRIDOTICSIRIATICS -ETENIRIDOTIOTICTIRIDOTOCTIOTICTIRIDS复制启动,以病毒卵巢肿瘤 - 类半胱氨酸去泛素酶(votu)结构域为由。投票域可能是导致TP从大型PPOLB多肽裂解的原因,并且在腺毒中被灭活,其中TP是单独的蛋白质。许多PLV和转囊编码了Polinton的独特衍生物 - 例如保留TP,Fotu和PPOLB聚合棕榈域的PPOLB,但缺乏外核酸酶域,而含有一个超家族1个旋转酶结构域。分析了在真核前肿瘤前胞菌中,对投票域的存在/不存在和将PPOLB用其他DNA聚合酶代替,使我们能够概述其起源和进化的完整情况。
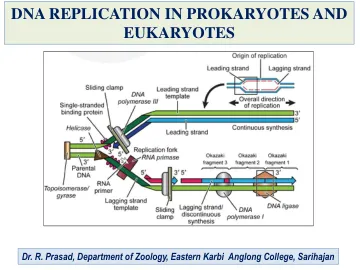
原核生物和真核生物中的DNA复制
DNA的复制始于在称为复制起源的位点放松双螺旋。在这些位点,碱基之间的氢键被损坏,并且成对底座分开。一对复制片段聚集在一起并连接非复制DNA的位置称为复制叉。在细菌染色体中,DNA复制总是从称为原点的特定位点开始。每个来源控制一个称为复制子的DNA单元的复制。细菌具有复制的单个特定起源

具有双果冻的真核DNA病毒的自然史
门前病毒(Kingdom Bamfordvirae,Realm varidnaviria)是多种病毒的广泛组合,其相对较短的双链DNA基因组(<50 kbp)产生了由双果冻 - 双果冻 - 卷胶卷蛋白构建的二十os虫。前肿瘤动物感染所有细胞结构域的宿主,证明其古老的起源,尤其是与真核生物的七个超级组中的六个有关。前肿瘤分子包括四个主要的病毒组,即Polinton,Polinton,例如病毒(PLV),病毒噬细胞和腺毒。我们使用蛋白质结构建模和分析来表明蛋白质的DNA聚合酶(PPOLBS),polins,病毒噬细胞和细胞质线性质粒涵盖了n-终末结构域与末端蛋白(TPS)的N-末端域同源物(TPS),例如原始prd1-涉及tpectiricotic andototic artectirIdotics和eukaryotic artirIdotics artirIdotic artirIdotic artineciridotics anden tectirifiridotic toNERIFIRIDICRIDOTICSIRIATICS -ETENIRIDOTIOTICTIRIDOTOCTIOTICTIRIDS复制启动,以病毒卵巢肿瘤 - 类半胱氨酸去泛素酶(votu)结构域为由。投票域可能是导致TP从大型PPOLB多肽裂解的原因,并且在腺毒中被灭活,其中TP是单独的蛋白质。许多PLV和转囊编码了Polinton的独特衍生物 - 例如保留TP,Fotu和PPOLB聚合棕榈域的PPOLB,但缺乏外核酸酶域,而含有一个超家族1个旋转酶结构域。分析了在真核前肿瘤前胞菌中,对投票域的存在/不存在和将PPOLB用其他DNA聚合酶代替,使我们能够概述其起源和进化的完整情况。

新西兰排放交易体系 (ETS) NZU 盈余建议
不得因报告内容或向第三方提供报告而对安永提出任何索赔或要求或任何诉讼或程序。安永将永远免于承担任何此类索赔、要求、诉讼或程序。在编制本报告时,安永已考虑并依赖客户和参与流程的其他利益相关者以及其他被认为可靠和准确的来源向我们提供的信息。安永并未被告知任何提供给它的信息或从公共来源获得的信息是虚假的,或任何重大信息被隐瞒。安永并不暗示,也不应被解释为安永已对提供给我们的任何信息执行了审计、验证或尽职调查程序。安永未独立验证任何此类信息,也不承担独立验证此类信息的任何责任或义务,也不对信息的准确性或完整性作出任何陈述。安永或其任何成员或员工均不承担任何责任,对于因向安永提供不正确信息而导致本报告中的错误而给任何人造成的任何损失或损害,亦不承担任何责任。