XiaoMi-AI文件搜索系统
World File Search System减数分裂

2 发展的生物和环境基础
人体中的大多数细胞通过称为有丝分裂的过程进行繁殖,在此过程中,DNA 自我复制,复制染色体,最终形成具有相同遗传物质的新细胞 (Sadler, 2018)。有丝分裂过程负责所有体细胞的复制。然而,性细胞以不同的方式繁殖,即通过减数分裂。首先,46 条染色体开始像有丝分裂一样复制自身。但在细胞完成分裂之前,会发生一个称为交叉的关键过程。染色体对对齐,DNA 片段交叉,从染色体对的一个成员移动到另一个成员,本质上是“混合”了 DNA。因此,交叉会产生独特的基因组合 (Sadler, 2018)。由此产生的细胞仅由 23 条单个未配对的染色体组成。这些细胞被称为配子,专门用于有性生殖:男性是精子,女性是卵子。卵子和精子在受精时结合,产生受精卵,即合子,它有 46 条染色体,形成 23 对,一半来自亲生母亲,一半来自亲生父亲。每个配子都有独特的遗传特征,据估计,个体可以产生数百万个遗传不同的配子(美国国家医学图书馆,2019 年)。

细菌CRISPR/CAS9系统作为所有健康问题的有希望的解决方案,并进步的生物工程
缩写:BP1,肿瘤抑制剂p53结合蛋白1; BRCA,乳腺癌抗原;汽车,嵌合抗原受体; CAS9,CRISPR相关蛋白9;级联,抗病毒防御的CRISPR综合体; CMR,CAS模块坡道(重复相关的神秘蛋白质); CMR III-B,多个亚基III型B CRISPR RNA-CAS蛋白; CPF1,Prevotella和Francisella1的CRISPR; CRISPR,定期间隔间隔室; Crrna,Crispr RNA; CSM III-A,多支亚基III-A CRISPR-CAS蛋白; dcas9/ sgrna-sg I,停用cas9/短指南RNA-Sybrr-green i; DNA-PK,DNA-蛋白K; DNA-PKC,DNA蛋白K催化亚基; DSB,双链断裂; ege,额外的基因元素; GRNA,导向RNA; HDR,同源性维修; IAP,碱性磷酸酶同工酶; MRE 11,减数分裂重组11; NHEJ,非同理结局加入; PAM,原始间隔者相邻基序; PD,程序性细胞死亡; RAD,重组酶A;代表,重复的外部回文; RPA,复制蛋白A; RT,逆转录酶; Sgrna,简短的指南RNA; SSB,单链断裂; tracrrna,反式激活CRISPR RNA; XLF,类似XRCC4的因子; XRCC 4,X射线修复交叉补充蛋白4; Yoyo-1,(恶唑黄色)

除草剂利谷隆对热带爪蟾大脑和睾丸 DNA 甲基化谱的雄性传递跨代效应
除草剂利谷隆可对非洲爪蟾(Xenopus Tropicalis)产生内分泌干扰作用,包括从未接触过该污染物的后代。这些影响跨代传递的机制有待进一步研究。在这里,我们研究了大脑和睾丸 DNA 甲基化谱的跨代改变,这些改变是从发育过程中接触到环境相关浓度利谷隆的祖父那里遗传下来的。简化代表性亚硫酸氢盐测序 (RRBS) 揭示了成年雄性 F2 代大脑 (3060 个 DMR) 和睾丸 (2551 个 DMR) 中的许多差异甲基化区域 (DMR)。大脑中参与生长激素 ( igfbp4 ) 和促甲状腺激素信号传导 ( dio1 和 tg ) 的关键基因存在差异甲基化,并与体型、体重、后肢长度和血糖水平的表型改变相关,表明这些甲基化变化可能是利谷隆跨代效应的潜在介质。睾丸 DMR 存在于精子发生、减数分裂和生殖细胞发育所必需的基因( piwil1 、 spo11 和 tdrd9 )中,其甲基化水平与每个曲细精管的生殖细胞巢数量相关,这是精子发生中断的终点。DMR 还存在于调节表观遗传景观的机制(包括 DNA 甲基化)的几个基因中
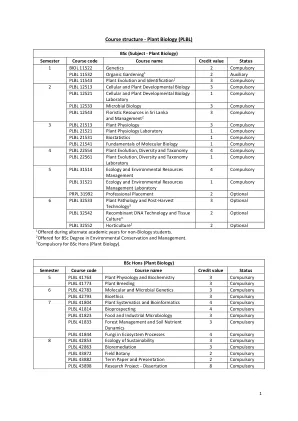
课程结构 - 植物生物学(PLBL)
Course unit contents - Plant Biology (PLBL) Semester 1 Course Code BIOL 11522 Course Name Genetics Credit Value 2 Core / Optional Core Pre-requisites GCE A/L Biology Co-requisites - Hourly Breakdown Theory Practical Independent Learning 30 hrs 15 hrs 55 hrs Course Aim / Intended Learning Outcomes : Upon successful completion of this course unit, the student should be able to, (i) describe the principles of inheritance, (ii)解释分子遗传学和(iii)在遗传学背景下解决基本问题所获得的知识。课程内容:审查孟德尔遗传学和孟德尔继承模式的扩展。等位基因变异的分子基础。链接和基因映射。具有农业和生物医学相关性的复杂性状的定量遗传学。遗传变异和种群遗传学应用的来源。遗传物质的分子组织。DNA复制和修复。核基因组,基因,基因表达和基因表达调节的简介:乳糖操纵子。人类基因组项目,选定遗传疾病的遗传和分子基础,基因检测和基因治疗的介绍。选定模型生物基因组的基本原理。分子生物学和遗传学的应用。实验室:显微镜,细胞分裂:有丝分裂和减数分裂,强硬韦恩伯格平衡和自然选择的演示,人类遗传教学 /学习方法:讲座,实验室会议和教程评估策略:持续评估和结束课程单位书面考试< / div> < / div> < / div> < / div> < / div> < / div>

核糖体 DNA 不稳定性是核型进化的潜在原因
核型是指基因组构成一组染色体的结构。物种间的核型差异预计会阻碍各种生物过程,如染色体分离和减数分裂染色体配对,从而可能导致不相容性。核型可以在近缘物种之间甚至同一物种的不同种群之间迅速变化。然而,人们对驱动核型进化的力量了解甚少。在这里,我们描述了从塞舌尔群岛分离出来的果蝇品系的独特核型。该品系丢失了 X 染色体上的核糖体 DNA (rDNA) 位点。由于 Y 染色体是唯一其他携带 rDNA 的染色体,所以所有雌性都携带至少一条 Y 染色体作为 rDNA 的来源。有趣的是,我们发现该品系还携带一条截短的 Y 染色体 (YS ),尽管它无法支持男性生育能力,但它在种群中稳定维持。我们的建模和细胞学分析表明,Y 染色体对雌性适应度的负面影响大于 YS 染色体。此外,我们生成了一个独立的菌株,该菌株缺乏 X rDNA,其核型为 XXY 雌性和 XY 雄性。该菌株迅速进化出多种核型:两个新的截短 Y 染色体(类似于 YS ),以及两个独立的 X 染色体融合,其中包含 Y 衍生的 rDNA 片段,从而消除了雌性对 Y 染色体的依赖。考虑到罗伯逊融合经常发生在人类的 rDNA 基因座上,我们提出 rDNA 基因座不稳定性可能是核型进化的驱动力之一。

植物无融合生殖的分子基础
植物的有性生殖是一个复杂且受到严格调控的过程,可产生新一代的散播体:有性种子。传统上,在创造新作物品种的过程中,有性生殖被用来分离或选择性地组装所需的基因和性状。然而,有性的利用也给植物育种带来了限制,包括种子成本高昂且方法耗时。在植物育种过程中,可以通过依次利用有性和无融合生殖来缓解大多数这些限制。无融合生殖是一种协同机制的结果,该机制利用性机制并以协调胚珠发育步骤的方式发挥作用,从而产生无性(克隆)种子。有性发育的改变涉及减数分裂、配子发生以及胚胎和胚乳形成中广泛表征的功能和解剖变化。无融合生殖植物的胚珠跳过减数分裂,形成未减数的雌配子体,其卵细胞发育成孤雌生殖胚胎,中央细胞可能与精子融合,也可能不融合,形成种子胚乳。因此,功能性无融合生殖至少涉及三个组成部分,即无融合生殖 + 孤雌生殖 + 胚乳发育,这些组成部分是从有性生殖改良而来的,必须在分子水平上进行协调,才能完成发育步骤并形成克隆种子。尽管最近在发现与无融合生殖样表型和克隆种子形成相关的特定基因方面取得了进展,但无融合生殖的分子基础和调控网络仍然未知。这是目前无融合生殖育种局限性的核心问题。本期特刊汇集了 12 篇围绕无融合生殖分子基础的不同主题的出版物,展示了最近在理解该性状的遗传调控方面取得的发现和进展,并讨论了无融合生殖的可能起源及其在植物中商业化应用的其他挑战。由于无融合生殖是一种基于有性生殖功能获得或丧失突变的现象的理论仍未得到解决,Barcaccia 等人 [ 1 ] 重新评估了被子植物无融合生殖的进化起源及其替代发育途径,并提出了系统发育和遗传证据,支持无融合生殖是从有性生殖进化而来的,是由于有性发育中关键参与者的分子破坏而导致的。此外,Schmidt [ 2 ] 概述了高等植物无融合生殖的分子方面,并清楚地解释了无融合生殖发育所涉及的调控复杂性,强调了 DNA 和 RNA 结合蛋白以及非编码 RNA 在通过表观遗传调控机制激活和抑制发育程序中的积极作用。同样,Ortiz 等人 [ 3 ] 在以 Paspalum spp. 为例的研究中总结了有关无融合生殖的大量信息。并详细介绍了该属无融合生殖发育的关键方面和所使用的各种遗传分析,包括基因组位点的分子表征、三个生殖候选基因( ORC3 、 QGJ 和 TGS1 )的功能表征以及进一步基于基因组的研究路线图。从不同的植物物种中获得了有关无融合生殖的进一步分子细节。Mateo de Arias 等人 [ 4 ] 使用遗传和细胞胚胎学分析结合应激处理对五个物种进行了研究,以提供大量证据支持多态性

单细胞转录组揭示斑马鱼卵巢的发育和功能
摘要:斑马鱼是一种成熟的研究生物,为我们理解脊椎动物组织和器官的发育做出了许多贡献,但我们对调节性腺发育、性别和生殖的基因的理解仍然存在重大差距。与许多器官(如大脑和心脏)在发育的最初几天内形成的发育不同,斑马鱼性腺直到幼虫阶段(受精后 ≥ 5 天)才开始形成。因此,正向遗传筛选已确定了极少数性腺发育所需的基因。此外,识别性腺中表达基因的大量 RNA 测序研究没有足够的分辨率来定义可能在这些器官的发育和功能中发挥重要作用的小细胞群。为了克服这些限制,我们使用单细胞 RNA 测序来确定从幼年斑马鱼卵巢中分离的细胞的转录组。这得到了 10,658 个生殖细胞和 14,431 个体细胞的图谱。我们的生殖细胞数据代表了从生殖系干细胞到早期减数分裂卵母细胞的所有发育阶段。我们的体细胞数据代表了所有已知的体细胞类型,包括卵泡细胞、卵泡膜细胞和卵巢基质细胞。进一步分析发现,在这些广义的细胞类型中,存在数量出乎意料的细胞亚群。为了进一步确定它们的功能意义,我们确定了这些细胞亚群在卵巢内的位置。最后,我们使用基因敲除实验来确定 foxl2l 和 wnt9b 分别对卵母细胞发育和性别决定和/或分化的作用。我们的结果揭示了斑马鱼卵巢发育和功能的新见解,转录组谱将为未来的研究提供宝贵的资源。

非同源末端连接是 CRISPR/Cas 的关键……
尽管成簇的规律间隔短回文重复序列 (CRISPR)/CRISPR 相关蛋白 (Cas) 介导的基因编辑已经彻底改变了生物学和植物育种,但大规模的可遗传植物染色体重组仍处于起步阶段。现在可以实现染色体内的重复和倒位,以及染色体之间的易位。随后,可以破坏或新建遗传连锁。此外,染色体上基因的顺序也可以改变。虽然自然染色体重组在减数分裂过程中通过同源重组发生,但 CRISPR/Cas 介导的染色体重排最好通过利用体细胞中的非同源末端连接 (NHEJ) 途径获得。NHEJ 可细分为经典 (cNHEJ) 和替代 NHEJ (aNHEJ) 途径,它们部分地以拮抗方式运作。 cNHEJ 通路不仅可以保护断裂的 DNA 末端免于降解,还可以抑制先前未连接的断裂末端的连接。因此,在没有 cNHEJ 的情况下,可以获得更多的倒位或易位,这可以归因于无限制地使用 aNHEJ 通路进行双链断裂 (DSB) 修复。与倒位或易位相反,短串联重复可以通过 Cas9 切口酶由成对的单链断裂产生。有趣的是,cNHEJ 通路对于这些类型的重复至关重要,而 aNHEJ 则是补丁插入所必需的,补丁插入也可以在 DSB 修复期间形成。由于染色体工程不仅在模式植物拟南芥 (Arabidopsis thaliana) 中实现,而且在作物玉米 (Zea mays) 中也实现,我们预计这项技术将很快改变育种过程。

最终生物学教学大纲2025 PDF
●什么是生活?生物多样性;需要分类;生命的三个领域;分类学和系统学;物种和分类层次结构的概念;二项式术语;研究分类法的工具 - 博物馆,动物园,草药,植物园。●五个王国分类:Monera的显着特征和分类; protista和真菌分为主要群体;地衣;病毒和病毒,将植物的显着特征和分类为主要群体,苔藓植物,孢子菌,裸子植物和被子植物;被子植物 - 分类为类,特征特征和示例,显着特征和动物 - 非对抗的分类,直至门水平,然后缔结级别。●动物和植物中的结构组织:形态和修饰;组织;解剖学和流动植物的不同部分的功能:根,茎,叶,渗透性 - cymose和camose和comemose,豆类,水果和种子,动物组织;昆虫(蟑螂)的不同系统(消化,循环,呼吸,神经和生殖)的形态,解剖学和功能。●细胞结构和功能:细胞理论和细胞作为生命的基本单位;原核和真核细胞的结构;植物细胞和动物细胞;细胞包膜,细胞膜,细胞壁;细胞细胞器结构和功能;内膜系统 - 肾上腺素网,高尔基体,溶酶体,液泡;线粒体,核糖体,质体,微生物;细胞骨架,纤毛,叶叶菌,中心元素(超微结构和功能);核核膜,染色质,核仁。●细胞分裂:细胞周期,有丝分裂,减数分裂及其意义。活细胞的化学成分:蛋白质,碳水化合物,脂质,核酸的生物分子结构和功能;酶类型,性质,酶作用。

实现 CRISPR/Cas 介导的基因组编辑的策略,无需转基因整合即可直接获得编辑植物
在过去的一个世纪里,随着植物遗传学理解的加深以及强大且易于使用的基因编辑工具的开发,人类传递精确作物基因型的能力发生了革命性的变化。植物转化技术已经很发达,可用于在某些作物和模式生物中制造转基因品种,但试剂输送和植物再生仍然是将基因编辑技术应用于大多数作物的关键瓶颈。生产转基因、基因改造 (GM) 品种的典型植物转化方案依赖于转基因、化学选择和组织培养。制造基因编辑 (GE) 品种的典型方案也使用转基因,即使这些转基因可能对最终的作物产品不利。在某些作物中,转基因通常在减数分裂期间通过杂交分离出来,因此这只是一个次要的问题。在其他作物中,特别是那些无性繁殖的作物、复杂的杂交种或世代时间长的作物,这种杂交是不切实际的或不可能的。本综述重点介绍了将 CRISPR/Cas 基因编辑试剂递送至可再生植物细胞并恢复已编辑植物而不产生不必要的转基因整合的各种策略。一些示例包括递送无 DNA 的基因编辑试剂(如核糖核蛋白或 mRNA)、依赖非整合 DNA 的试剂表达、使用病毒或纳米颗粒等新型递送机制、使用非常规选择方法避免转基因整合和/或完全避免组织培养。这些方法正在迅速发展,并已使作物科学家能够利用 CRISPR 基因编辑工具的精确性。