XiaoMi-AI文件搜索系统
World File Search System功能域
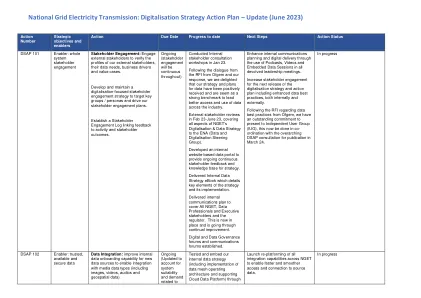
数字化战略行动计划 – 更新版(2023 年 6 月)
更新的 NGET 数据域设计已完成并实施。这主要集中在资产、客户、网络、财务和人员等主要数据域上。我们已认识到存在许多子域或跨域,例如投资、监管和供应商,并已将其与功能域所有权和产品开发联系起来。

Pioneer Factor 改进了基于 CRISPR 的 C-To-G 和 C-To-T 碱基编辑
真核生物中的碱基编辑事件需要兼容的染色质环境,但关于染色质因子如何影响编辑效率或窗口的研究很少。通过设计与各种先驱因子融合的BE(碱基编辑器),作者发现SOX2显著提高了GBE和CBE的编辑效率。SoxN-GBE(SOX2-NH3-GBE)提高了原型间隔物整体胞嘧啶的编辑效率,而SoxM-GBE/CBE(SOX2-Middle-GBE/CBE)则能够在PAM-近端胞嘧啶处实现更高的碱基编辑。通过分离SOX2的功能域,构建了SadN-GBE(SOX2激活域-NH3-GBE)以获得更高的编辑效率,而SadM-CBE则具有更宽的编辑窗口。通过 DNase I 试验,还证明了编辑效率的提高很可能与 SAD 诱导染色质可及性有关。最后,使用 SadM-CBE 在原癌基因 MYC 中引入终止密码子,该位点以前很少被高效编辑。在这项工作中,通过融合先锋因子或其功能域构建了一类新的先锋 BE,它在真核生物中表现出更高的编辑效率或更宽的编辑窗口。

引用:Fırat C、Öztürk A、Dağcı İ、Solak K、Ünver Y NanoScript:一种基于纳米粒子的新型基因调控工具及其优点和缺点。E
转录因子 (TF) 是一种蛋白质,它通过与特定 DNA 序列结合,通过与基因组中的特定调控元件相互作用来激活或抑制基因表达,从而充当基因表达的关键调节器。TF 通常具有多个功能域,这些功能域有助于其调节功能。这些功能域基本上由三个域组成:核定位信号 (NLS) 域、DNA 结合域 (DBD) 和激活域 (AD)。通过这些域的协调相互作用,TF 响应细胞内的各种内部和外部信号来调节基因表达。TF 复杂机制的缺陷与越来越多的人类疾病有关。因此,基于 TF 的基因调控研究被认为是许多生物应用的有前途的方法。在这种情况下,研究人员旨在使用一种称为 NanoScript 的基于纳米粒子的平台来模拟 TF 的结构和功能特性。NanoScript 的作用类似于天然 TF,可实现精确的基因调控和细胞重编程,并为控制和有针对性地操纵基因表达提供了新的可能性。 NanoScript 的主要目标是以非病毒方式在转录水平上调节基因表达。NanoScript 可以通过与内源 DNA 相互作用并启动转录活性来激活特定基因,作为基因操作和细胞重编程的蛋白质替代合成结构。该平台由于其可调组件(纳米粒子和表面组件)和有效调节基因表达的能力,在干细胞生物学、癌症治疗和细胞重编程领域具有多种应用潜力。然而,NanoScript 也有一些局限性,例如可能与脱靶基因相互作用。本研究讨论了 NanoScript 在基因调控领域的当前研究和技术,以及该技术的优势和挑战。

pdx1突变继发于突变的两例新生儿糖尿病的临床和分子描述
对基因或基因产物的有害影响(保护,进化,剪接影响等))美国医学遗传与基因组学学院(ACMG)的标准在15个预测因子中分析了将其归类为致病性。 2。 根据PM1, pdx1 c.571a> g(p.lys191glu),处于杂合性的状态,被分类为具有不确定意义的变体(位于突变热点和/或关键且良好的功能域(例如 酶的活性位点)没有良性变化); pm2(不存在对照(或很少经常隐性)); PP3(多种计算证据支持对基因或基因产物的有害影响(保护,进化,剪接影响等)))在15个预测因子中分析了将其归类为致病性。2。pdx1 c.571a> g(p.lys191glu),处于杂合性的状态,被分类为具有不确定意义的变体(位于突变热点和/或关键且良好的功能域(例如酶的活性位点)没有良性变化); pm2(不存在对照(或很少经常隐性)); PP3(多种计算证据支持对基因或基因产物的有害影响(保护,进化,剪接影响等))标准,在15个预测因子中进行了分析,其中14个将其归类为致病性,并具有中等效应。数据库中未报告此变体。PTF1A基因未报告致病性变异。

构建,管理和分析质粒库的最终指南
例如,要研究基因在疾病模型中的作用,您可以构建由基因的各种功能域或具有缺失域和靶向突变的变体组成的质粒文库。以这种方式,您可以破译该基因的生物学作用,基因的功能结构域以及与该基因功能相关的关键氨基酸残基。此外,您可以根据正在从事的不同项目以及用于制备质粒DNA的过程(研究与临床等级材料或简单的DNA Prep与哺乳动物细胞的无内毒素质粒准备)创建质粒文库。

一个用于基因组数据组合分析的灵活工具包
在这里,我们介绍了TrackPlot,这是一个Python软件包,用于通过可编程和基于互动的Web方法生成出版物质量可视化。与生成实地的程序的现有范围相比,TrackPlot提供了一个多功能平台,可在各种来源中视觉解释基因组数据,包括具有功能域映射,同种型,同种型的基因注释,而没有通过SCRNA-SECRED和长期访问的范围以及杂色的范围,以及任何杂色的访问范围,并提供了透明度的范围,以及杂色的范围。符合主要期刊要求的输出文件。TrackPlot软件包是一种开源软件,可以在Bioconda(https://anaconda.org/bioconda/trackplot)上免费获得,Docker(https://hub.docker.com/r/r/r/r/ygidtu/trackplot) (https://github.com/ygidtu/trackplot),还提供了用于本地部署的内置Web服务器。

CEP55 抑制剂
有丝分裂时膜脱落的部位。在中体中,CEP55 通过 EABR 结构域与 ESCRT-I 亚基成员 TSG101(肿瘤易感基因 101)和 ALIX(ALG-2 相互作用蛋白 X)直接相互作用,募集运输机制所需的内体分选复合物 (ESCRT) 成分,然后募集 ESCRT-III 完成胞质分裂。7,8 在多种与癌症患者预后不良相关的肿瘤中均发现 CEP55 的过表达。CEP55 的上调促进小鼠的基因组不稳定性和肿瘤形成。体外研究发现,CEP55 过表达通过 PI3K/Akt 信号通路诱导不受控制的细胞增殖。9 相反,CEP55 失调与细胞分裂不完全和异常有关。 10,11 这种抗癌策略的预期结果是抑制 CEP55 活性导致细胞异常分裂,随后癌细胞死亡。因此,本研究提供了有关 CEP55 基因的全面信息,包括其表达、功能域、重要的磷酸化位点以及异常的不良临床病理特征

利用 CRISPR–Cas9 平铺筛选对人类有丝分裂基因进行功能解析
人类蛋白质编码基因的身份众所周知,但我们对其分子功能和域结构的深入了解仍然受到基于同源性的预测和专注于全基因耗竭的实验方法的缺陷的限制。为了弥补这一知识空白,我们开发了一种方法,利用 CRISPR - Cas9 诱导的蛋白质编码基因突变来先验地识别序列水平的功能区域。作为一个测试案例,我们将这种方法应用于 48 个人类有丝分裂基因,揭示了细胞增殖所需的数百个区域,包括实验表征的区域、基于同源性预测的区域和新区域。我们验证了 15 个区域的筛选结果,包括 Mad1 的 387 - 402 个氨基酸,这些氨基酸以前未被表征,但有助于 Mad1 着丝粒定位和染色体分离保真度。总之,我们证明基于 CRISPR - Cas9 的平铺诱变可以从头识别蛋白质编码基因中的关键功能域,从而阐明功能突变体的分离并允许跨人类蛋白质组的功能注释。

CRISPR/CAS9介导的基因靶向的通用系统使用植物中多合一的载体
同源重组介导的基因组编辑,也称为基因靶向(GT),是一种必不可少的技术,允许对目标序列进行精确的修改,包括引入点突变,报告基因的敲入和/或交换功能域。然而,由于其低频,很难建立可以广泛应用于大量植物物种的GT方法。我们开发了一种简单且通用的定期间隔短的短粒子重复序列(CRISPR)/CRISPR相关的蛋白9(CAS9)介导的DNA双重链突破(DSB)诱导的GT系统,使用包含CRIS CRIS/CAS9表达构造的多一对矢量,可供选择的标记和GT Donor donor donor template。该系统启用了具有不可选择的特征的目标点突变,以大米和烟草中的几个靶基因。可以精确地使用该系统评估内源性靶基因的GT频率,因此我们研究了用RAD51刺激化合物1(RS-1)处理对DSB诱导GT频率的治疗的影响。GT频率略有但始终如一,通过RS-1处理在两个目标植物中都得到了改善。

Caf
摘要组蛋白伴侣染色质组装因子1(CAF-1)沉积了两个新生的组蛋白H3/H4二聚体在新复制的DNA上,形成了核小体的中心核心,称为Tortasome。CAF-1如何确保有足够的空间来组装四面体。Caf-1的赖氨酸/谷氨酸/精氨酸(KER)区域的结构和生物物理表征揭示了具有前所未有的DNA结合特性的128-Å单α-螺旋(SAH)基序。不同的KER序列特征和SAH的长度驱动Caf-1对于四长长的DNA的选择性,并促进发芽酵母中的功能。在体内,KER与CAF-1中的DNA结合的有翼螺旋结构域合作,以克服DNA损伤敏感性并保持基因表达的沉默。我们建议KER SAH将CAF-1中的功能域与结构精度联系起来,在染色质组装过程中充当DNA结合间隔元件。